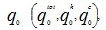Тренинг по типу мышечных волокон и их строение. Какие волокна мышц обладают более быстрой сократительной способностью
В чём отличия медлнных (ST) и быстрых (FT-A и FT-B) мышечных волокон
Типы мышц, их строение являются той причиной, по которой один атлет может бежать быстрее и наращивать мышечную массу легче других, а другой — способен бежать длительное время без признаков утомления.
Такую разницу создают процессы, которые происходят в мышечной ткани. Важно понимать их, например, для того чтобы выстроить правильную тренировочную программу, которая подойдет для конкретно выбранного спортсмена.
почему сокращаются мышцы?
- Пресинаптическое окончание.
- Сарколемма.
- Синаптический пузырек.
- Никотиновый ацетилхолиновый рецептор.
- Митохондрия
К волокнам скелетных мышц подходят толстые нервные волокна, которые отходят от передних отделов спинного мозга. После попадания в мышцу, каждое нервное волокно делится и снабжает своими разветвлениями до нескольких сотен мышечных волокон.
Соединение нерва и мышечного волокна образует так называемый синапс, или нервно-мышечное соединение (причем на каждом мышечном волокне формируется только один такой синапс). Под влиянием нервного сигнала возникает так называемый потенциал действия, который распространяется от спинного мозга по нервам к мышце и синапсу.
То, как скелетная мускулатура будет адаптироваться к повторяющимся стимулам, в большей степени зависит от внутренних характеристик самой мышцы. Именно типы мышечных волокон вносят наибольший вклад в возможность выполнения спортсменом той или иной тренировочной программы.
Типы мышечных волокон
У людей выделяют три типа мышечных волокон:
- Медленносокращающиеся (slow-twitch, ST или I тип) волокна характеризуются медленным временем сокращения, а также большой сопротивляемостью усталости. В своей структуре эти волокна имеют маленький мотонейрон и диаметр нервного волокна, высокую плотность митохондрий и капилляров, большое содержание миоглобина.
Этот тип волокон имеет небольшое количество креатин фосфата — высокоэнергетического субстрата, необходимого для быстрого, взрывного движения, — а значит, эти волокна не способны сокращаться быстро.
Функционально, ST-волокна используются при аэробной активности, не требующей большого приложения силы, например — ходьба и поддержание позы. Большая часть повседневной активности задействует именно ST-волокна.
- Быстросокращающиеся (fast-twitch, FT или II тип) волокна характеризуются быстрым сокращением и низкой сопротивляемостью усталости. Разница в скорости сокращения, от которой произошло разделение волокон, может быть отчасти объяснена скоростью выделения кальция из саркоплазматического ретикулума (место в клетке, где хранится кальций), а также активностью фермента, который расщепляет АТФ внутри головки миозина (один из сократительных белков). Обе эти характеристики быстрее и в большей степени присутствуют в FT-волокнах.
Сами FT-волокна делятся на два типа: тип А (FT-A или IIA тип) и тип В (FT-B или IIB тип).
- Волокна FT-A имеют умеренную сопротивляемость усталости и представляют собой переходный тип между медленносокращающимися волокнами и волокнами типа FT -B. Функционально они используются при длительной анаэробной активности с относительно большой продукцией силы, например, беге на 400 метров.
- С другой стороны, быстросокращающиеся волокна типа B, очень чувствительны к усталости и используются для коротких анаэробных нагрузок с большой продукцией силы, такие как: спринтерские забеги, бег с барьерами, прыжки. Эти волокна также способны продуцировать больше энергии, чем ST волокна.
Краткие характеристики волокон приведены в таблице ниже.
| Тип волокна | Медленносокр.(ST) | Быстросокр. тип А(FT—A) | Быстросокр. тип B(FT—b) |
| Время сокращения | Медленное | Быстрое | Очень быстрое |
| Размер мотонейрона | Маленький | Большой | Очень большой |
| Сопротивляемость усталости | Высокая | Средняя | Низкая |
| Активность | Аэробная | Длительная анаэробная | Короткая анаэробная |
| Продукция силы | Низкая | Высокая | Очень высокая |
| Плотность митохондрий | Высокая | Высокая | Низкая |
| Плотность капилляров | Высокая | Средняя | Низкая |
| Окислительная способность | Высокая | Высокая | Низкая |
| Гликолитическая емкость | Низкая | Высокая | Высокая |
| Преимущественный запас топлива | Триглицериды (жиры) | Креатинфосфат, гликоген | Креатинфосфат, гликоген |
При конкретной скорости движения, количество продуцируемой силы определяется типом вовлеченного в движение мышечного волокна. Во время динамического сокращения, когда волокно укорачивается или удлиняется, быстросокращающиеся волокна продуцируют больше силы, чем медленносокращающиеся волокна.
В условиях, когда длина мышечного волокна не изменяется при сокращении, ST-волокна продуцируют столько же силы, как и FT-волокна. Разница в продукции силы наблюдается только при активном сокращении, когда изменяется длина волокна.
При конкретной скорости движения, сила, продуцируемая мышцей, возрастает с повышением содержания FT-волокон, и наоборот, при конкретной продукции силы, скорость повышается при увеличении количества FT-волокон.
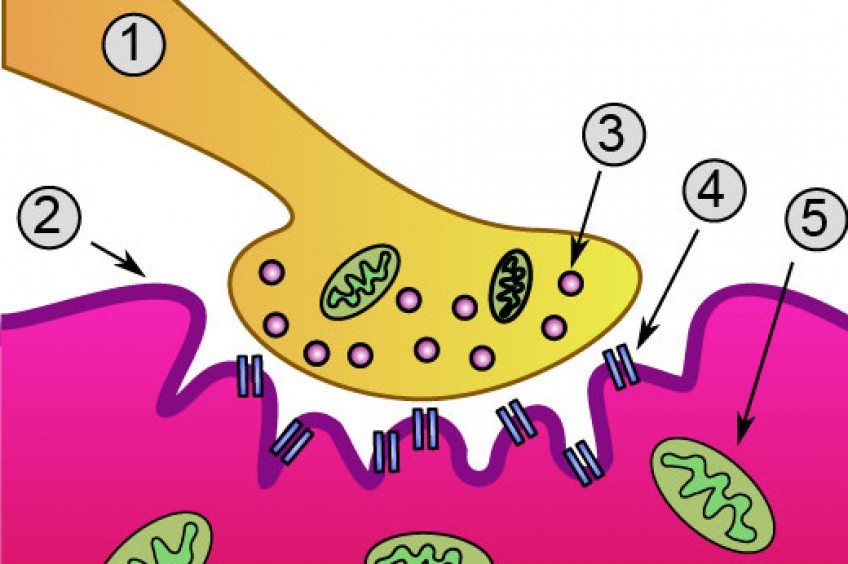
Существует большой разброс в процентном соотношении волокон у атлетов. Например, хорошо известно, что участвующие в соревнованиях на выносливость имеют большее содержание медленносокращающихся волокон, тогда как спринтеры и прыгуны имеют больше быстросокращающихся волокон.
Больший процент FT-волокон у спринтеров позволяет им продуцировать большую силу и мощность, чем у атлетов с повышенным содержанием ST-волокон.
Разница в составе мышц у атлетов подняла вопрос о том, наследуется ли такой состав генетически или он может быть изменен с помощью тренировок. Исследования, проведенные на близнецах, показали, что в основном состав мышц и процент содержания в них разных типов волокон определяется генетически.
Тем не менее, есть ряд данных, доказывающих, что как структура, так и метаболическая емкость мышечных волокон может изменяться в ответ на различные типы тренировок.
Вовлечение мышечных волокон в работу
Мышца продуцирует силу путем вовлечения так называемых моторных единиц — группы мышечных волокон, которую «обслуживает» одно двигательное нервное окончание. Во время произвольного изометрического и концентрического сокращения, обычный порядок вовлечения моторных единиц контролируется их размерами — это состояние известно под названием «принцип размера».
Маленькие моторные единицы, содержащие медленносокращающиеся мышечные волокна, имеют наименьший порог активизации, т.е. для их активизации достаточно самого слабого стимула, поэтому они вовлекаются первыми. Потребность в выработке большей силы удовлетворяется вовлечением более крупных моторных единиц.
Самые большие моторные единицы, содержащие быстросокращающиеся волокна типа B имеют наивысший порог активизации, и поэтому вовлекаются последними. Вне зависимости от интенсивности работы, первыми в нее вовлекаются медленносокращающиеся волокна.
Если интенсивность работы низкая, то медленносокращающиеся волокна остаются единственными, вовлеченными в нее. Если интенсивность работы высокая, например, подъем тяжелого веса, или интервальная работа на стадионе, первыми вовлекаются медленносокращающиеся волокна, затем подключаются быстросокращающиеся волокна типа A, а затем, при необходимости, волокна типа B.
Существуют данные о том, что принцип размера может быть нарушен или даже полностью изменен во время некоторых типов движений, особенно тех, которые содержат эксцентрические (с удлинением мышцы) компоненты. При этом быстросокращающиеся волокна могут активизироваться раньше медленносокращающихся.
Определение типа волокон
Поскольку единственным способом напрямую определить состав мышечного волокна у атлета является проведение биопсии мышцы (прямой метод), то некоторые исследования попробовали определить состав мышечного волокна непрямым методом, путем выявления взаимосвязей между различными свойствами типа волокна и состава волокон мышц.
Интересные данные, полученные в этих исследованиях, показывают значимую взаимосвязь между содержанием быстросокращающихся волокон и мышечной силой или мощностью.
Непрямой метод, который можно использовать для определения состава волокон мышц, состоит в определении максимального веса, который спортсмен может поднять всего один раз.
После этого производится максимальное количество повторов с весом в 80% от максимального. Если общее количество повторений меньше семи, скорее всего мышцы более чем на 50% состоят из FT-волокон. Если же удается сделать двенадцать и более повторов, скорее всего, мышца более чем на 50% состоит из ST-волокон. Если число повторений между 7 и 12 — скорее всего, мышцы состоят поровну из FT и ST-волокон.
Поскольку подъем веса вовлекает большое количество групп, этот метод не работает при определении состава изолированных мышц, а только мышечных групп.
Для определения состава волокон отдельной мышцы может потребоваться игольчатая биопсия интересующей мышцы. Другим непрямым методом, который можно использовать, является участие в различных соревнованиях. Доминантные волокна можно выявить, исходя из успеха в определенных соревнованиях, что позволит в дальнейшем развивать именно эти способности мышц.
Применение на практике
Пропорция типов волокон в мышцах будет влиять не на то, какой вес вы сможете поднять, какое количество повторений вы сможете сделать в интервальной работе, а на конечный результат — повышение силы/мощности мышц или выносливости.
Например, атлет, в мышцах которого большое содержание быстросокращающихся волокон, будет неспособен выполнить такое же количество повторений с весом, как атлет, в мышцах которого содержатся преимущественно медленносокращающиеся волокна.
Таким образом, атлет с FT-волокнами никогда не достигнет той мышечной выносливости, которая будет у атлета с ST-волокнами. Аналогично, атлет с большей пропорцией ST-волокон в мышцах не сможет поднять такой же вес, или пробежать интервалы так же быстро, как и атлет с большей пропорцией FT-волокон в мышцах. Следовательно, атлет с ST-волокнами не будет таким же сильным и мощным, как атлет с FT-волокнами.
Однако необходимо помнить, что даже внутри группы спринтеров или бегунов на длинные дистанции будет большой разброс по типам волокон в мышцах. Не все спринтеры имеют одинаковый процент FT-волокон, не все бегуны на длинные дистанции имеют одинаковый процент ST-волокон. Поэтому, одни спринтеры могут сделать работу 12х200 м, тогда как другие устанут после 8 повторов.
В зависимости от типа волокна и быстроты наступления утомления (из-за большего количества FT-волокон) необходимо решить, нужно ли больше отдыхать между интервалами для того, чтобы закончить работу, или необходимо уменьшить количество интервалов и увеличить скорость в серии.
Тренировка FT-волокон мышцы для выносливости не увеличит количество ST-волокон, а тренировка ST-волокон для силы и мощности не приведет к увеличению количества FT-волокон.
При соответствующем тренинге, FT-B волокна могут принять на себя некоторую часть выносливости, характерную для FT-BA волокон, а FT-A волокна могут принять на себя некоторую часть силы и мощности, характерной для FT-B волокон.
Однако, не существует полной взаимозаменяемости волокон. FT-волокна не могут стать ST-волокнами, и наоборот. Другими словами, то, с каким процентным соотношением волокон родился человек, с таким он будет жить и тренироваться.
Несмотря на то, что тип волокон не может быть изменен с одного на другой, тренировки могут изменить ту площадь, которую занимает определенный тип волокон в мышце. Другими словами, может произойти выборочное увеличение волокон, путем воздействия на них тренировками.
Например, у атлета в мышце может быть соотношение FT/ST-волокон 50/50, но поскольку площадь поперечного сечения FT волокон обычно больше, чем у ST-волокон, 65% площади мышцы могут занимать быстросокращающиеся, а 35% — медленносокращающиеся волокна.
При тренировках с отягощениями для повышения силы мышц, соотношение FT/ST-волокон останется таким же — 50/50, однако изменится площадь поперечного сечения, занимаемая двумя типами волокон. Это произойдет, потому что ST-волокна атрофируются, а FT-волокна гипертрофируются.
В зависимости от интенсивности тренировки, площадь мышцы может состоять на 75% из FT-волокон, и на 25% из ST-волокон. Эти изменения повлекут за собой повышение силы, но уменьшение выносливости.

Кроме этого, поскольку масса FT-волокон больше, чем ST-волокон, атлет будет набирать массу, если измерить окружности мышц. Напротив, если атлет тренируется для повышения выносливости, FT-волокна атрофируются, а ST-волокна гипертрофируются, вызывая увеличение площади поперечного сечения ST-волокон.
Площадь мышцы, изначально состоявшая на 65% из FT и на 35% из ST-волокон, может измениться под влиянием тренировок, и соотношение будет 50% на 50%. Кроме того, из-за того, что масса ST-волокон меньше, чем FT, наряду с повышением выносливости, произойдет снижение силы, а также потеря некоторой части мышечной массы.
Известный факт: если необходим прирост силы мышц, нужно тренироваться с тяжелыми весами и небольшим количеством повторов.
Этот режим тренировок приводит к рекрутированию FT-B волокон, которые могут развивать большее усилие, чем ST или FT-A волокна. Гипертрофия развивается только в перегруженной мышце, поэтому во время тренировки будет происходить рекрутмент FT-B волокон и их дальнейшая гипертрофия.
Тренировки с низкой или умеренной интенсивностью не всегда приводят к рекрутменту FT-B волокон, следовательно, для вовлечения этих волокон интенсивность должна быть высокой.
Выводы
Необходимо помнить, что для максимального результата необходимо тренироваться в соответствии со своей генетической предрасположенностью или на основании того, на каких соревнованиях вы показываете лучшие результаты.
Например, для атлета с преобладанием медленносокращающихся волокон, большую пользу принесет увеличение километража и тренировки с небольшими весами и большим количеством повторов.
С другой стороны, те атлеты, у которых преобладают быстросокращающиеся волокна, извлекут больше пользы из «спринтерских» методов тренировок и тренировок с тяжелыми весами и небольшим количеством повторов.
Было доказано, что длинные пробежки способствуют развитию медленносокращающихся волокон, улучшая их аэробные качества, тренируя их устойчивость к утомлению. Темповые тренировки влияют на ST и FT-A волокна, среди прочих эффектов улучшая работу этих типов волокон в связке.
Интервальная работа вовлекает FT-A и FT-B волокна, тренируя их взаимодействие и улучшая нейромышечную координацию. Скоростная работа: набегания, спринт в горку, короткие спринты на дорожке стадиона позволяют по максимуму нагрузить FT-B волокна.
Возможно, это не принесет большой пользы тем, кто бегает марафон, но увеличение силы позволит вам бежать более плавно и эффективно.
Автор статьи: доцент кафедры анестезиологии и реаниматологии СЗГМУ им. Мечникова Евгений Суборов
trail-run.ru
Основные типы и характеристики мышечных волокон
В этой статье давайте рассмотрим основные типы и характеристики мышечных волокон,а также выясним какое соотношение мышечных волокон преобладает у человека по половому признаку и виду физической деятельности.
Классификация скелетных мышц основана на различиях в тяжёлых цепях молекул миозина, которые преобладают в их составе. Чтобы понять о чём идёт речь и представить общую картину, взглянем на строение мышцы.

Всего существует 3 типа мышечных волокон, но основные принято выделять два типа: быстрые физические (или тип II) и медленные физические (или тип I). Эти типы волокон различаются по своим метаболическим и сократительным характеристикам. Рассмотрим подробную характеристику каждого типа.
 Быстрые физические волокна способны развивать быстрое и сильное сокращение. Это происходит благодаря ряду особенностей их метаболизма, в частности более высокой скорости выброса ионов кальция, более высокой активности миозиновой АТФазы и хорошо развитому саркоплазматическому ретикулуму. Скорость сокращения и развиваемое усилие быстрых физических волокон в 3—5 раз выше аналогичных показателей медленных физических волокон. Быстрые физические волокна используют как энергетический источник преимущественно глюкозу крови и собственные запасы гликогена, и поэтому участвуют главным образом в двигательной активности анаэробного типа, например при выполнении подачи в волейболе, теннисе или на тренировочных занятиях силовой направленности. Быстрые физические волокна могут быть разделены на две основные группы: тип A и тип B. Быстрые и медленные волокна могут содержать в своём составе миозиновые молекулы только с одним типом тяжёлых цепей, либо включать различные типы молекул миозина с преобладанием одного из них. Волокна типа А считают волокнами промежуточного типа, поскольку они обладают примерно в равной степени способностью производить энергию аэробным и анаэробным путём, поэтому их ещё называют быстрыми волокнами окислительного типа. Волокна типа Б проявляют наиболее высокий анаэробный потенциал, поэтому их называют быстрыми волокнами с гликолитическим типом окисления.
Быстрые физические волокна способны развивать быстрое и сильное сокращение. Это происходит благодаря ряду особенностей их метаболизма, в частности более высокой скорости выброса ионов кальция, более высокой активности миозиновой АТФазы и хорошо развитому саркоплазматическому ретикулуму. Скорость сокращения и развиваемое усилие быстрых физических волокон в 3—5 раз выше аналогичных показателей медленных физических волокон. Быстрые физические волокна используют как энергетический источник преимущественно глюкозу крови и собственные запасы гликогена, и поэтому участвуют главным образом в двигательной активности анаэробного типа, например при выполнении подачи в волейболе, теннисе или на тренировочных занятиях силовой направленности. Быстрые физические волокна могут быть разделены на две основные группы: тип A и тип B. Быстрые и медленные волокна могут содержать в своём составе миозиновые молекулы только с одним типом тяжёлых цепей, либо включать различные типы молекул миозина с преобладанием одного из них. Волокна типа А считают волокнами промежуточного типа, поскольку они обладают примерно в равной степени способностью производить энергию аэробным и анаэробным путём, поэтому их ещё называют быстрыми волокнами окислительного типа. Волокна типа Б проявляют наиболее высокий анаэробный потенциал, поэтому их называют быстрыми волокнами с гликолитическим типом окисления.
 Медленные физические волокна, или волокна типа I, обычно участвуют в выполнении продолжительной работы, связанной с потреблением энергии, образованной аэробным путем, — это занятия степ-аэробикой, выполнение упражнений в воде, бег на длинные дистанции, занятия на велотренажёрах в постоянном темпе. Этот тип волокон характеризуется как устойчивый к утомлению, тогда как быстрые физические волокна не способны к продолжительной нагрузке. Медленные волокна отличаются менее развитым саркоплазматическим ретикулумом, вследствие чего снижается скорость выброса кальция и АТФазная активность миозина, что в свою очередь замедляет гидролиз АТФ. Кроме того, медленные волокна отличаются более низкой способностью к гликолизу. Однако волокна типа I содержат значительное количество митохондрий, ферменты которых повышают их способность к выполнению работы с аэробной продукцией энергии. Из-за высокой доли аэробного метаболизма в энергообеспечении сокращения и низкой скорости сокращения волокна типа I часто называют ещё медленными волокнами окислительного типа. Волокна этого типа обычно хорошо снабжаются кровью благодаря развитой системе кровеносных капилляров, что является структурной и функциональной адаптацией к их значительной потребности в кислороде.
Медленные физические волокна, или волокна типа I, обычно участвуют в выполнении продолжительной работы, связанной с потреблением энергии, образованной аэробным путем, — это занятия степ-аэробикой, выполнение упражнений в воде, бег на длинные дистанции, занятия на велотренажёрах в постоянном темпе. Этот тип волокон характеризуется как устойчивый к утомлению, тогда как быстрые физические волокна не способны к продолжительной нагрузке. Медленные волокна отличаются менее развитым саркоплазматическим ретикулумом, вследствие чего снижается скорость выброса кальция и АТФазная активность миозина, что в свою очередь замедляет гидролиз АТФ. Кроме того, медленные волокна отличаются более низкой способностью к гликолизу. Однако волокна типа I содержат значительное количество митохондрий, ферменты которых повышают их способность к выполнению работы с аэробной продукцией энергии. Из-за высокой доли аэробного метаболизма в энергообеспечении сокращения и низкой скорости сокращения волокна типа I часто называют ещё медленными волокнами окислительного типа. Волокна этого типа обычно хорошо снабжаются кровью благодаря развитой системе кровеносных капилляров, что является структурной и функциональной адаптацией к их значительной потребности в кислороде.
Соотношение мышечных волокон у человека
Интересно отметить, что в мышцах верхних и нижних конечностей одного человека соотношение доли быстрых и медленных волокон обычно почти одинаково. Единственным исключением является камбаловидная мышца — сгибатель стопы, участвующий в ходьбе и видах деятельности, связанных с переносом тяжести, для чего необходимы, прежде всего, медленные волокна. У основной массы мужчин, женщин и детей мышцы конечностей состоят на 45—55 % из медленных мышечных волокон. Волокна типов А и Б также представлены примерно в равной мере.
Половые различия в распределении различных типов мышечных волокон практически отсутствуют, они проявляются лишь в абсолютных размерах мышц. Однако если речь идёт о профессиональных спортсменах высокого класса, здесь обнаруживаются заметные отличия. У спринтеров проявляется тенденция к преобладанию в мышцах ног быстрых волокон, в то время как у спортсменов, которым необходима высокая аэробная выносливость, наблюдается доминирование медленных волокон. У легкоатлетов — бегунов на средние дистанции доля медленных и быстрых волокон часто почти одинакова. Преобладание определённого типа мышечных волокон является лишь одним показателем, определяющим успех спортсмена, и не может использоваться как универсальное средство прогноза результативности спортсмена.
Соотношение быстрых и медленных волокон в мышцах человека, очевидно, определяется уже в первые годы его жизни, т. е. является генетически предопределённым и практически не изменяется до самой старости. По мере старения у людей проявляется тенденция к утрате быстрых волокон в результате возрастных изменений и недостаточной двигательной активности.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
street-sport.com
опорно двигательный аппарат быстрые и медленные мышечные волокна

Чтобы успешно заниматься физическими упражнениями с отягощениями и на тренажерах, необходимо иметь четкое представление об опорно двигательном аппарате человека.
Опорой всех человеческих тканей и органов является скелет, состоящий из множества костей. Подвижные соединения в костном скелете — их насчитывается до 230 — называются суставами. Концы сочленяющихся костей плотно охвачены соединительной оболочкой, называемой суставной сумкой.
Основную роль в укреплении суставов играют связки — прочные и упругие тяжи из соединительной ткани. Они, срастаясь с соединительной сумкой, укрепляют ее. Большое значение в укреплении суставов имеют сухожилия, прикрепляющиеся к костям. Для разнообразия движений в некоторых суставах имеются особые пластинки или диски из соединительно-тканного волокнистого вещества. Выделяемая в полость сустава внутренними слоями тканей суставной сумки вязкая жидкость (синовия) снижает трение между контактирующими поверхностями костей. Основные ключевые движения в суставах это:
- а) сгибание,
- б) разгибание,
- в) приведение,
- г) отведение,
- д) ротация (вращение),
- е) круговые движения.
Благодаря силовым упражнениям повышается крепкость суставов, они становятся более подвижными. Однако при запредельной (чрезмерной) нагрузке и значительном превышении степени свободы вероятны травмы — вывихи, порой даже с разрывом тканей и кровеносных сосудов.
Все движения человек выполняет благодаря сократительной деятельности более шестисот скелетных мышц. Различают два вида мускулатуры — гладкую, сокращающуюся помимо воли (желудок, стенки кровеносных сосудов), и поперечнополосатую, перемещающую тело в пространстве, за счет управляемого человеком мышечного сокращения. В состав поперечнополосатой мышцы входят тонкие нити белка актина и толстые нити — миозина, которые, объединяясь, образуют саркомеры — элементарные двигательные единицы, где происходит преобразование химической энергии в механическую, вызывая движение человека.
Имеется предположение, что сократительный процесс мышцы возникает в результате взаимного проникновения нитей актина и миозина. В связи с этим энергетический уровень саркомера зависит от положения этих нитей в нем. Объединяясь в группы, саркомеры образуют более тысячи тонких нитей — фибрилл, из которых и состоит мышечное волокно. Волокна образуют мышечные пучки, а те, объединяясь, — саму мышцу. Сократительные волокна мышцы оканчиваются у соединительной ткани, которая переходит в сухожилие и переносит напряжение при сокращении. Соединительная ткань обладает высокой прочностью.
Виды мышц
В зависимости от внешнего вида мышцы получили следующие названия:
- длинные,
- короткие,
- широкие,
- кольцеобразные.
Почти все широкие мышцы расположены на туловище, длинные — в основном на конечностях, короткие — между отдельно взятыми позвонками. Визуально длинные мышцы похожи на веретено. Средняя часть такой мышцы называется «брюшком», начало зовётся «головкой», а второй конец (который более длинный) — «хвостом».
Некоторые мышцы имеют несколько головок или посередине перетягиваются сухожилистыми образованиями, разделяющими их на несколько частей. Сухожилия мышцы прикрепляются к всевозможным шероховатостям, бугристостям и различным выступам костей, прочно вплетаясь в надкостницу и даже частично проникая вглубь костного вещества, а в некоторых случаях к суставной сумке, фасции или коже.
Механика движений человека
При сокращении мышца перемещает кости, выступающие в роли рычагов, в суставах. Она, относительно немного укорачиваясь, развивает довольно большие усилия. Поэтому в опорно двигательном аппарате человека обычно имеют место костные рычаги с проигрышем силы при работе мышцы, но с выигрышем пути приложения этой силы. Величина момента силы зависит от угла, под которым сила действует на рычаг. Наибольший эффект достигается, когда сила действует под прямым углом к рычагу.
С изменением угла сгибания в локтевом суставе от 0 до 100° происходит увеличение плеча силы в среднем от 11,5 до 44,5 мм, или проще - в четыре раза, причем максимум внешней силы достигается при угле 90°. Однако в действительности момент внешней силы человека значительно меньше максимального из-за приложения силы мышцы к кости не под прямым углом.
Различные бугры и выступы на костях скелета, а также сесамовидные косточки (например, коленная чашечка) способствуют более рациональному воздействию мышцы на костные рычаги. Мышцы, вызывающие при сокращении движение звеньев тела только в одном суставе, называются односуставными, а прикрепленные своими концами одновременно к костям я отдельным частям скелета и приводящие к изменению углов сразу во многих суставах — многосуставными.
При выполнении суставного движения за счет сокращения определенных групп мышц-синергистов — всегда (за исключением наличия противодействия внешних сил) можно вернуть движущееся звено в исходное положение благодаря наличию мышц-антагонистов.
Сила мышцы зависит от анатомического строения. Выделяют мышцы, имеющие перистое строение, веретенообразное с параллельными волокнами. Установлено, что мышцы перистого строения короткие и приспособлены к развитию напряжения большой силы (например, икроножная), а мышцы с параллельными и веретенообразными волокнами более длинные и обеспечивают быстрые, ловкие и размашистые движения (портняжная, двуглавая плеча).
Быстрые и медленные мышечные волокна
Сила мышц тем больше, на сколько больше их площадь поперечного сечения, а величина сокращения тем выше, чем длиннее мышечные волокна. Некоторые мышцы могут укорачиваться до трети - половины исходной длины. В мышцах имеются быстрые и медленные мышечные волокна. Первые, представленные в основном в перистых мышцах, например в икроножной, сокращаются быстрее медленных при прочих равных условиях. Сокращение также зависит от внешней нагрузки, от деятельности центральной нервной системы и от силы самой мышцы.
Связь величины силы мышцы с поперечником обусловлена количеством ее составляющих волокон. К примеру, одиночное поперечнополосатое волокно может развивать напряжение 0,1 - 0,2 г.
Анатомия движений
Сократительная способность характеризуется абсолютной силой, развиваемой всей мышцей, приходящейся на 1 см2 поперечного сечения (физиологический поперечник). Это позволяет сравнивать силу различных мышц, независимо от их величины. Например, абсолютная сила а) икроножной мышцы в сумме с камбаловидной равна 6,24, б) двуглавой плеча —11,4, в) трехглавой плеча — 16,8, г) плечевой — 12,1 кг/см2. Физиологический поперечник у некоторых мышц значительно превосходит анатомический поперечник.
Мышца сокращается благодаря импульсу, поступающему из центральной нервной системы (на одиночный импульс — одиночное сокращение). Чем выше нагрузка, тем больше скрытый период момента поступления импульса до момента сокращения. Величина этого сокращения зависит от приложенной внешней нагрузки: чем она больше, тем в меньшей степени укорачивается мышца.
Достигнув максимума сокращения после одиночного раздражения, мышца снова расслабляется и удлиняется до исходного уровня. Но это происходит не мгновенно, а в течение некоего времени. Поэтому если, не дав мышце полностью расслабиться, повторить раздражение, она опять сократится, но еще быстрее и мощнее, чем в первый раз. При частых импульсах раздражения происходит слияние одиночных сокращений в одно, называемое тетанусом.
В спортивных движениях или при обычной мышечной деятельности всегда присутствует тетаническое сокращение скелетной мускулатуры, причем тем более высокое, чем сильнее и чаще происходит импульсация из центральной нервной системы.
В неработающей мышце всегда есть некоторое напряжение, и она слегка сокращена за счет поступающих слабых импульсов. Это обстоятельство во многом и определяет рельеф мускулатуры, что особенно выражено у атлетически сложенных спортсменов.
Каждому состоянию мышцы соответствует ее определенная длина. Если нет препятствий со стороны внешних факторов, то с изменением своего физиологического состояния мышца стремится принять длину, соответствующую этому состоянию. В случае, когда за счет внешних условий длина и физиологическое состояние мышцы не соответствуют друг другу (если длина мышцы больше ее длины в ненагруженном состоянии), она деформирована относительно собственной длины, т. е. растянута. Учитывая упругие свойства мышцы, можно говорить о наличии потенциальной энергии упругой деформации, благодаря которой при изменении внешних условий может совершаться работа по перемещению окружающих костных рычагов и связанных с ними других тел.
Третий закон Ньютона
Мышечная тяга рождается в результате непосредственного взаимодействия нашего двигательного аппарата с всевозможными внешними объектами. Разновидность мышечной работы определяется характером этого взаимодействия — соотношением между внутренними и внешними силами. Если основной момент сил группы мышц превышает момент сил, противодействующей тяге, они осуществляют преодолевающую работу, а в противном случае — уступающую. При этом, когда моменты сил мышечной тяги равны сопротивлению, мы имеем дело с удерживающим видом мышечной работы. В положении основной стойки мышцы ног работают в статическом режиме, во время приседания — в уступающем, а при выпрямлении ног — в преодолевающем.
Таким образом, физической работе статического или динамического характера всегда предшествует изменение потенциальной энергии упругой деформации мышц.
Каждая мышца в организме выполняет строго определенную двигательную функцию. Рассмотрим самые основные из них:
Мышцы плечевого пояса.
- Грудино-ключично-сосцевидная мышца крепится к рукоятке грудины, внутреннему концу ключицы и к височной кости черепа (так называемому сосцевидному отростку). При одновременном сокращении правой и левой мышцы голова человека наклоняется вперед; при одностороннем сокращении происходит вращение и наклон головы соответственно в сторону задействованной мышцы.
- Дельтовидная мышца является мощной поверхностной мышцей, которая имеет крепление к дельтовидной бугристости, находящейся в верхнем участке плечевой кости. В зависимости от остальных креплений и функций она подразделяется на ключичную, плечевую и лопаточную, причем все три части способны к самостоятельному сокращению. Передняя часть мышцы отводит руку вперед и поворачивает внутрь; средняя часть производит отведение руки в сторону, отведение вперед и вверх; а вот задняя - отводит руку вверх, назад и вращает наружу.
- Малая круглая мышца крепится к нижнему и верхнему краям лопатки и к большому бугру на плечевой кости. Обеспечивает вращение плеча наружу и приведение руки.
- Большая круглая тянется от нижнего угла лопатки до гребня малого бугра плечевой кости. Участвует в тяге плеча вниз и назад и в его вращении.
- Двуглавая мышца плеча (бицепс) имеет две головки и один хвост. Она берет свое начало в ямке плечевого сустава и так называемого клювовидного отростка и закрепляется к лучевой кости. Бицепс сгибает плечо, а также предплечье в локтевом суставе, участвует во вращении предплечье наружу.
- Трехглавая мышца плеча (трицепс) имеет 3-ри головки: длинная имеет начало от лопатки, внутренняя и внешняя головки - от плечевой кости. В итоге все эти 3-ри головки сходятся к единому сухожилию, закрепленному на локтевой отросток локтевой кости. Мышца разгибает предплечье.
- Мышцы предплечий делятся на мышцы передней и задней групп. Мышцы передней группы сгибают кисть и пальцы в кулак, производят вращение предплечья внутрь, сгибают в локтевом суставе. Мускулатура задней группы производит разгибание кисти и пальцев, а также вращает предплечье наружу, разгибает его.
Мускулатура груди.
- Большая грудная мышца проходит поверхностно и обладает треугольной формой. Начинаясь от внешнего участка ключицы, грудины, конкретнее от хрящей 2—7-го рёбер, она крепится к плечевой кости – точнее к гребню ее большого бугорка. Участвует в движениях приведения руки к торсу, а также вращает ее внутрь.
- Малая грудная мышца имеет веерообразную форму и расположена глубже большой. При сокращении оттягивает лопатку по направлению вперед и вниз.
Мышцы спины.
- Трапециевидная группа размещается в верхней трети спины. Ее верхняя часть поднимает лопатку, нижняя — опускает, а средняя — приближает к позвоночнику. В результате сокращения мышцы лопатка приводится к средней линии. Верхняя ее часть в значительной степени предопределяет внешний контур шеи, так как имеет свое начало непосредственно в области шеи и распространяется до 12-ого грудного позвонка.
- Широчайшая мышца спины охватывает нижне-боковой отдел спины человека и, поднимаясь вверх, крепится к гребню плечевой кости – опять же малого ее бугорка. Эта мышца тянет плечом руку назад, а также одновременно вращает её внутрь. Еще она приводит нижний угол лопатки спины к грудной клетке.
- Глубокие мышцы спины расположены по обеим бокам у самого позвоночника практически по всей его длине и образуют длинный разгибатель позвоночника.
Мускулатура живота.
- Наружная косая мышца торса широким пластом проходит снаружи и сверху вниз. Начинается зубцами от 8-ми нижних ребер. Спереди и внизу перетекает в широкое плоское сухожилие, называемое апоневроз. Косые мышцы торса обеспечивают наклонные движения позвоночника в всевозможные стороны и его повороты вправо и влево.
- Прямая мышца живота лежит снаружи от средней линии и идет продольно сверху вниз. Сухожильными образованиями делится на 4 части и, следовательно, имеет четыре брюшка. Участвует в сгибании туловища вперед.
Мышцы ног.
- Большая и малая ягодичные мышцы. Большая производит вращение бедра кнаружи, с одновременным разгибанием его. Малая — отводит бедро.
- Четырехглавая мышца нижней конечности (квадрицепс) — разгибает нашу голень в коленном суставе, а также сгибает бедро.
- Двуглавая мышца бедра расположена на задней его поверхности у наружного края. Она выполняет сгибание голени в коленном суставе, производит разгибание в тазобедренном суставе, поворачивает голень наружу.
- Сгибание голени осуществляется также с помощью полусухожильной, полуперепончатой и стройной мышц задней поверхности бедра.
Важно понимать, что без теории – нет практики. Поэтому, только досконально изучив, как устроен наш опорно двигательный аппарат, можно добиться выдающихся достижений в фитнесе и бодибилдинге. Только четко понимая, как работает наше тело, можно приступать к его строительству. Так что не ленитесь лишний раз заглядывать в теорию. Чем больше Вы знаете – тем меньше ошибок Вы будете делать и меньше времени потратите, а это – дорогого стоит…
- < Назад
- Вперёд >
www.fitness-bodybuilding.ru
Некоторые общие положения
Спорт  Некоторые общие положения
Некоторые общие положения
просмотров - 93
Контрольные вопросы
1. Гомеостаз это....
2. Организм это....
3. Почему кости детей более эластичны и упруги?
4. Посредством чего кости скелета соединяются между собой?
5. К какому виду мускулатуры относятся скелетные мышцы?
6. Сколько мышц насчитывается у человека?
7. Какие волокна мышц обладают более быстрой сократительной способностью?
8. На что расщепляется гликоген при анаэробных процессах образования энергии?
9. Что образуется при окислении углеводов и жиров?
10. Какой процесс энергообразования обладает большими возможностями во времени?
11. Какую функцию осуществляют клетки крови - эритроциты?
12. Какой пульс считается нормальным для здорового взрослого человека?
13. Какое кровяное давление является нормой у здорового человека в возрасте 18-40 лет?
14. Какое воздействие оказывают систематические занятия физическими упражнениями на дыхательную мускулатуру?
15. Какой процент глюкозы, образуемой печенью, потребляется головным мозгом?
16. Укажите наиболее эффективную форму отдыха при умственном труде.
17. От чего защищает организм такое функциональное состояние, как утомление?
18. До какого уровня восстанавливаются энергетические ресурсы в восстановительном периоде после физической нагрузки?
19. Когда лучше тренироваться, учитывая биологические ритмы?
20. К чему приводит пониженная двигательная активность?
21. Какова продолжительность работы в зоне умеренной мощности?
22. Как отличаются расходы энергии в покое у тренированных и нетренированных людей?
23. На что указывает урежение пульса в покое (брадикардия) у тренированных людей?
24. В каких видах спорта наблюдается тесная связь между максимальным потреблением кислорода (МПК) и тренированностью?
25. Какова норма потребления белков в день для взрослого человека?
26. Что является главным источником энергии в организме?
27. Когда преимущественно используются жиры как источник энергии при физической деятельности?
28. Каково основное значение витаминов для организма?
29. Сколько калорий расходует в течение рабочего дня (8-10 ч) мужчина, занимающийся умственным и физическим трудом?
30. Какое количество энергии крайне важно затрачивать ежедневно для нормальной жизнедеятельности?
31. Какова причина "гравитационного шока"?
32. Физические упражнения какого характера оказывают наиболее эффективное воздействие на сердечно-сосудистую систему?
33. Какова причина так называемой "мертвой точки"?
34. Как можно ослабить проявление "мертвой точки"?
35. Какие меры способствуют качественной готовности студентов к активной учебной работе?
Глава 3. ОСНОВЫ ЗДОРОВОГО ОБРАЗА ЖИЗНИ
Профилактика заболеваний и формирование здорового образа жизни населения, особенно молодежи, являются сегодня актуальной проблемой.
Среди современной молодежи можно выделить две взаимно противоположные группы: одна резко ограничивает свою двигательную активность и игнорирует средства физического воспитания, а другая одержима большими спортивными результатами и стремлением использовать для этого максимальные тренировочные и соревновательные нагрузки. И то и другое не способствует укреплению здоровья, гармоничному развитию человека и готовности к разнообразной деятельности. По этой причине крайне важно творческое осмысление теоретических и практических основ существующей системы физического воспитания с учетом крайне важности укрепления здоровья молодежи.
В ХIХ веке немецкий философ А. Шопенгауэр говорил: «Вообще 9/10 нашего счастья основано на здоровье. При нем все становится источником наслаждения». Известно также крылатое высказывание английского натуралиста Джона Рея, жившего в 17-м веке: "Здоровье дороже богатства".
Формирование здорового образа жизни у молодежи имеет исключительное значение. Молодые люди составляют до 30% населения земного шара. В молодости (16-29 лет) закладывается основа трудовых, нравственных позиций, а также здоровья человека. Что же такое здоровье?
По определению, принятому Всемирной организацией здравоохранения (ВОЗ), здоровье - ϶ᴛᴏ состояние полного физического, душевного и социального благополучия, а не только отсутствие болезни или физических дефектов.
По мнению Г.А. Апанасенко феномен жизни обеспечивается типовыми специализированными структурами, деятельность которых реализуется постоянной циркуляцией потоков пластических веществ, энергии и информации внутри системы, а также между ней и окружающей средой. На основании этого он дает следующее определение здоровью: «Здоровье – динамическое состояние человека, ĸᴏᴛᴏᴩᴏᴇ определяется резервами механизмов самоорганизации его системы (устойчивостью к воздействию патогенных факторов и способностью компенсировать патологический процесс), характеризуется энергетическим, пластическим и информационным (регуляторным) обеспечением процессов самоорганизации, а также служит основой проявления биологических (выживаемость-сохранение особи, репродукция-продолжение рода) и социальных функций".
Сегодня принято выделять следующие компоненты здоровья.
1. Соматическое – текущее состояние органов и систем органов человеческого организма.
2. Физическое – уровень развития и функциональных возможностей органов и систем организма. Основа физического здоровья - ϶ᴛᴏ морфологические и функциональные резервы клеток, тканей, органов и систем органов, обеспечивающие приспособление организма к воздействию различных факторов.
3. Психическое – состояние психической сферы человека. Основу психического здоровья составляет состояние общего душевного комфорта͵ обеспечивающее адекватную регуляцию поведения.
4. Сексуальное – комплекс соматических, эмоциональных, интеллектуальных и социальных аспектов сексуального существования человека, позитивно обогащающих личность, повышающих коммуникабельность человека и его способность к любви.
5. Нравственное – комплекс характеристик мотивационной и потребностно-информационной основы жизнедеятельности человека. Основу нравственного компонента здоровья человека определяет система ценностей, установок и мотивов поведения индивида в социальной среде.
В обобщенном и несколько упрощенном виде критериями здоровья являются: для соматического и физического здоровья – я могу; для психического здоровья – я хочу; для нравственного здоровья – я должен.
Читайте также
1. Определенность высказываний. Высказывания по поводу деятельности, которые квалифицируются рассказчиком как возможные или вероятные, учитываются, если они подпадают под ту или иную категорию. Например: Возможно, он только что работал. 2. Альтернативность... [читать подробенее]
1. Определенность высказываний. Высказывания по поводу деятельности, которые квалифицируются рассказчиком как возможные или вероятные, учитываются, если они подпадают под ту или иную категорию. Например: Возможно, он только что работал. 2. Альтернативность... [читать подробенее]
Контрольные вопросы 1. Гомеостаз это.... 2. Организм это.... 3. Почему кости детей более эластичны и упруги? 4. Посредством чего кости скелета соединяются между собой? 5. К какому виду мускулатуры относятся скелетные мышцы? 6. Сколько мышц насчитывается у человека? 7.... [читать подробенее]
oplib.ru
Механизм мышечного сокращения — SportWiki энциклопедия
Нервно-мышечная реакция на силовую тренировку[править]
Структура мышц[править]
Мышца - это комплексная структура, отвечающая за движение. Мышцы состоят из саркомеров, которые содержат определенное сочетание фибриллярных белков - миозина (толстые нити) и актина (тонкие нити), которые играют важную роль в мышечных сокращениях. Таким образом, саркомер - это сократительный элемент мышечного волокна, состоящий из миозиновых и актиновых белковых нитей.
Помимо этого, способность мышцы сокращаться и прилагать силу зависит конкретно от ее вида, площади поперечного сечения, а также длины и количества волокон внутри мышцы. Число волокон определяется генетикой, и на него невозможно повлиять с помощью тренировок; однако тренировки в состоянии изменить другие переменные. Например, число и толщина миозиновых нитей увеличивается посредством упорных тренировок с максимальной силовой нагрузкой. Увеличение толщины мышечных нитей увеличивает размер мышцы и силу сокращений.
Человеческое тело состоит из различных типов мышечных волокон, подразделяющихся на группы, и каждая группа относится к одной двигательной единице. В общем и целом в нашем организме имеются тысячи двигательных единиц, в которых находятся десятки тысяч мышечных волокон. Каждая двигательная единица содержит сотни или тысячи мышечных волокон, пребывающих в покое до тех пор, пока им не нужно действовать. Двигательная единица управляет совокупностью волокон и направляет их действия по закону «все или ничего». Этот закон означает, что при раздражении двигательной единицы импульс, направляемый в ее мышечные волокна, либо распространяется полностью - таким образом раздражая всю совокупность волокон, - либо не распространяется вообще.
Разные двигательные единицы реагируют на разные нагрузки при тренировках. Например, выполнение жима лежа с 60% повторного максимума задействует определенную совокупность двигательных единиц, тогда как более крупные двигательные единицы ожидают более высокой нагрузки. Поскольку последовательное задействование двигательных единиц зависит от нагрузки, необходимо разрабатывать специальные программы, чтобы активизировать и адаптировать основные группы двигательных единиц и мышечных волокон, играющих доминирующую роль в избранном виде спорта. К примеру, в тренировках для спринта на короткую дистанцию и легкоатлетических дисциплин (таких как толкание ядра) следует использовать тяжелые нагрузки, чтобы способствовать развитию силы, необходимой для оптимизации скорости и взрывных действий.
Мышечные волокна выполняют разные биохимические (метаболические) функции; выражаясь конкретнее, одни лучше приспособлены с физиологической точки зрения к работе в анаэробных условиях, а другие лучше работают в аэробных условиях. Волокна, которые используют кислород для выработки энергии, называются аэробными, тип I, красными или медленными. Волокна, которым кислород не требуется, называются анаэробными, тип II, белыми или быстрыми. Быстрые мышечные волокна, в свою очередь, делятся на подтипы IIА и IIХ (иногда называемые IIВ, хотя у людей тип IIВ практически не встречается[1]).
Медленные и быстрые волокна существуют примерно в равной пропорции. Однако в зависимости от их функций, в некоторых группах мышц (например, подколенные сухожилия, бицепсы) содержится больше быстрых волокон, тогда как в других (например, в камбаловидной мышце) содержится больше медленных волокон. В таблице 2.1 мы сравниваем характеристики быстрых и медленных волокон.
Сравнение быстрых и медленных волокон
| МЕДЛЕННЫЕ ВОЛОКНА | БЫСТРЫЕ ВОЛОКНА |
| Красные, тип I, аэробные | Белые, тип II, анаэробные |
| • Медленно устают • Нервная клетка меньше - иннервирует от 10 до 180 мышечных волокон • Развивают долгие, продолжительные сокращения • Применяются для развития выносливости • Активизируются во время низко- и высокоинтенсивной деятельности | • Быстро устают • Большая нервная клетка - иннервирует от 300 до 500 (или более) мышечных волокон • Развивают короткие, сильные сокращения • Применяются для развития скорости и силы • Активизируются только во время высокоинтенсивной деятельности |
Тренировки могут влиять на эти характеристики. Датские ученые Андерсен и Аагаард[2][3][4][5][6] в своих исследованиях показывают, что при объемных нагрузках или лактатных по природе тренировках волокна IIХ приобретают характеристики волокон IIА. То есть богатая миозином цепочка этих волокон становится более медленной и более эффективно справляется с лактатной деятельностью. Эти изменения можно повернуть вспять, снижая тренировочную нагрузку (тейперинг), в результате чего волокна IIХ возвращаются к изначальным характеристикам наиболее быстрых волокон[3]. Силовые тренировки также увеличивают размер волокон, благодаря чему вырабатывается больше силы.
Сокращение быстрой двигательной единицы более быстрое и мощное, чем сокращение медленной двигательной единицы. В результате пропорция быстрых волокон, как правило, выше в организме успешных спортсменов, занимающихся скоростно-силовыми видами спорта, но они также быстрее утомляются. Спортсмены с более высоким скоплением медленных волокон, напротив, обычно преуспевают в видах спорта на выносливость, поскольку они могут выполнять нагрузки низкой интенсивности в течение более продолжительного времени.
Активизация мышечных волокон происходит по принципу величины, известному также как принцип Хеннемана[7], согласно которому двигательные единицы и мышечные волокна активизируются начиная с меньшей в сторону большей. Активация всегда начинается с медленных волокон. При низкой или умеренно интенсивной нагрузке активируются медленные волокна и выполняют большую часть работы. При сильной нагрузке сначала сокращаются медленные волокна, затем в процесс вовлекаются быстрые волокна. При повторениях до отказа с умеренной нагрузкой двигательные единицы, состоящие из быстрых волокон, постепенно активизируются, чтобы поддерживать выработку силы, тогда как ранее задействованные двигательные единицы утомляются (см. рис. 1).
 рис. 1. Последовательная активизация двигательных единиц в подходе упражнений до концентрического отказа
рис. 1. Последовательная активизация двигательных единиц в подходе упражнений до концентрического отказа В распределении типов мышечных волокон у спортсменов, занимающихся разными видами спорта, могут наблюдаться различия. Это иллюстрируют рис. 2 и 2.3, представляющие общий процент содержания быстрых и медленных мышечных волокон у спортсменов в избранных видах спорта. Например, существенная разница между спринтерами и марафонцами четко дает понять, что успех в некоторых видах спорта хотя бы частично определяется генетическим составом мышечных волокон спортсмена.
 рис. 2. Распределение типов волокон у мужчин в разных видах спорта. Обратите внимание на преобладание медленных волокон у спортсменов, занимающихся аэробными видами спорта, и на преобладание быстрых волокон у спортсменов, занимающихся скоростно-силовыми видами спорта
рис. 2. Распределение типов волокон у мужчин в разных видах спорта. Обратите внимание на преобладание медленных волокон у спортсменов, занимающихся аэробными видами спорта, и на преобладание быстрых волокон у спортсменов, занимающихся скоростно-силовыми видами спорта Следовательно, пиковая мощность, вырабатываемая спортсменами, также имеет отношение к распределению типов волокон - чем выше процент быстрых волокон, тем большую мощность развивает спортсмен. Процент быстрых волокон также имеет отношение к скорости: чем выше скорость спортсмена, тем выше процент имеющихся у него быстрых волокон. Из таких людей получаются превосходные спринтеры и прыгуны, а подобный природный талант следует направлять в русло скоростно-силовых видов спорта. Попытка тренировать их, скажем, для бега на дистанцию означает трату таланта; в таких дисциплинах их ждет лишь средний успех, тогда как из них могут выйти отличные спринтеры, бейсболисты или футболисты (на этом список скоростносиловых видов спорта не кончается).
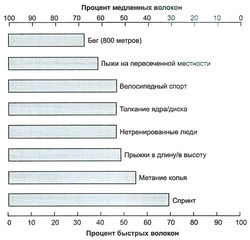 рис. 3. Распределение типов волокон у женщин в разных видах спорта
рис. 3. Распределение типов волокон у женщин в разных видах спорта Механизм мышечных сокращений[править]
Как мы описывали раньше, мышечные сокращения происходят в результате цепочки событий с участием белковых нитей - миозина и актина. В миозиновых нитях содержатся поперечные мостики - крошечные перемычки, выступающие вбок по направлению к актиновым нитям. Возбуждение, приводящее к сокращениям, стимулирует все волокно, создавая химические изменения, позволяющие актиновым нитям соединяться с миозиновыми поперечными мостиками. Связывание миозина с актином посредством поперечных мостиков высвобождает энергию, из-за чего поперечные мостики поворачиваются, таким образом подтягивая или совершая скользящее движение, связывающее миозиновые нити с актиновыми. Это скользящее движение вызывает мышечное сокращение, которое вырабатывает силу.
Чтобы визуализировать это иначе, вообразите гребную лодку. Весла представляют собой миозиновые нити, а воды - актиновые. Когда весла ударяются о воду, лодка с силой тянется вперед - и чем больше в воде весел, чем выше физическая сила гребца, тем больше вырабатываемая сила. Увеличение количества и толщины миозиновых нитей таким же образом повышает выработку силы.
Описанная ранее теория скользящих нитей дает понять, как работают мышцы, чтобы выработать силу. Эта теория включает в себя механизмы, способствующие эффективным мышечным сокращениям. Например, освобождение запаса эластичной энергии и рефлекторная адаптация играют ключевую роль в оптимизации спортивной работоспособности, но подобная адаптация происходит только тогда, когда в процессе тренировки происходит правильная стимуляция. Например, способность спортсмена использовать запас энергии для того, чтобы прыгать выше или толкать ядро дальше, оптимизируется посредством взрывных движений, как те, которые используются в плиометрическом тренинге. Однако мышечные компоненты - как, например, эластичные компоненты (сюда входят сухожилия, мышечные волокна и поперечные мостики) - не могут осуществлять эффективную транспортировку энергии, если спортсмен не укрепляет параллельные эластичные компоненты (напр., связки) и коллагеновые структуры (обеспечивающие стабильность и предохраняющие от травм). Если телу нужно выдерживать силы и воздействия, которым спортсмен подвергается, чтобы оптимизировать эластичные качества мышц, анатомическая адаптация должна предшествовать силовому тренингу.
Рефлекс - это непроизвольное мышечное сокращение, вызванное внешним стимулом[8]. Два основных компонента контроля рефлексов - это мышечные веретена и нервносухожильное веретено. Мышечные веретена реагируют на величину и скорость мышечного растяжения[9], тогда как нервно-сухожильное веретено (которое находится в местах соединения мышечных волокон с сухожильными пучками [8]) реагирует на мышечное напряжение. Когда в мышцах развивается высокая степень напряжения или растяжения, мышечные веретена и нервно-сухожильное веретено непроизвольно расслабляют мышцу, чтобы защитить ее от повреждения и травмы.
При пресечении этих ингибиторных реакций повышается спортивная работоспособность. Единственный способ добиться этого - адаптировать организм к более высокой степени напряжения, что повышает порог активизации рефлексов. Этой адаптации можно добиться посредством силового тренинга с использованием постепенно утяжеляющейся нагрузки (до 90 процентов повторного максимума или даже выше), таким образом вынуждая нервно-мышечную систему выдерживать более высокое напряжение, постоянно задействуя большее число быстрых волокон. В быстрых волокнах вырабатывается больше белка, что способствует увеличению силы.
Все спортивные движения выполняются по двигательной модели, которая называется циклом растяжение - сокращение и характеризуется тремя основными типами сокращения: эксцентрическим (удлинение), изометрическим (статичное положение) и концентрическим (сокращение). Например, волейболист, который быстро приседает и сразу подпрыгивает, чтобы блокировать атакующий удар, выполнил весь цикл растяжение - сокращение. То же касается и спортсмена, который опускает штангу на грудь и быстро выполняет взрывное движение, вытягивая руки. Чтобы полноценно пользоваться физиологическими качествами цикла растяжение - сокращение, мышца должна быстро переходить от удлинения к сокращению[10] (Schmidtble-icher, 1992).
Мышечный потенциал оптимизируется, когда активизируются все сложные факторы, влияющие на цикл растяжение - сокращение. Их влияние можно использовать для улучшения спортивных показателей только тогда, когда нервно-мышечная система стратегически стимулируется в правильной последовательности. Именно для достижения этой цели периодизация тренировки силы основывает планирование этапов на физиологической базе выбранного вида спорта. После составления эргогенного профиля (оценки вклада энергетических систем) выбранного вида спорта нужно пошагово распланировать этапы тренировки, чтобы перенести положительную нервно-мышечную адаптацию на практические показатели деятельности человека. Таким образом, понимание прикладной человеческой физиологии и установление цели в конце каждого этапа помогают тренерам и спортсменам интегрировать физиологические принципы в конкретную спортивную тренировку.
Повторим: скелетно-мышечная система тела - это сочетание костей, прикрепляемых друг к другу с помощью связок в области суставов. Пересекающие эти суставы мышцы дают силу для движения тела. Однако скелетные мышцы не сокращаются независимо друг от друга. Движения, выполняемые вокруг сустава, производятся несколькими мышцами, каждая из которых выполняет определенную роль, как уже было упомянуто выше.
Агонисты - или синергисты - это мышцы, которые взаимодействуют друг с другом при выполнении движения. В большинстве случаев, особенно если речь идет об умелом и опытном спортсмене, мышцы-антагонисты расслабляются, облегчая движение. Поскольку взаимодействие мышц группы агонистов и антагонистов напрямую влияет на спортивные движения, неправильное взаимодействие между этими группами может привести к порывистому или скованному движению. Следовательно, гладкость мышечного сокращения можно улучшить, если сосредоточиться на расслаблении антагонистов.
По этой причине одновременное сокращение (одновременная активизация мышц-агонистов и антагонистов, чтобы стабилизировать сустав) рекомендуется только на ранних стадиях реабилитации после травмы. Здоровому же спортсмену, особенно если он занимается силовыми видами спорта, не нужно выполнять упражнения (например, на нестабильной поверхности), вызывающие одновременные сокращения. К примеру, одной из основных характеристик элитных спринтеров является очень низкая миоэлектрическая активность мышц-антагонистов в каждой фазе цикла шага[11].
Первичные мышцы в первую очередь отвечают за суставное действие, которое является частью объемного силового движения или технической способности. Например, во время флексии локтя (сгибание бицепса) первичной мышцей является двуглавая мышца, тогда как трехглавая мышца (трицепс) выступает в роли антагониста и должна быть расслаблена, чтобы обеспечить беспрепятственное действие. В дополнение к этому стабилизаторы, или фиксаторы (обычно это меньшие мышцы), сокращаются изометрически, чтобы закрепить кость так, чтобы у первичных мышц была прочная база, откуда начинать натяжение. Мышцы других конечностей также могут принимать в этом участие, выступая в роли стабилизаторов, позволяющих первичным мышцам выполнять необходимые движения. Например, когда дзюдоист тянет соперника на себя, удерживая его за дзюдоги, мышцы его спины, ног и живота сокращаются изометрически, чтобы обеспечить стабильное основание для действия локтевых сгибателей (бицепсов), плечевых разгибателей (задние дельты) и лопаточных аддукторов и депрессоров (трапециевидная мышца и широчайшая мышца спины).
Механика мышечных сокращений[править]
Если мышцу стимулировать коротким электрическим импульсом, спустя небольшой латентный период происходит ее сокращение. Такое сокращение называется «одиночное сокращение мышцы». Одиночное мышечное сокращение длится около 10-50 мс, причем оно достигает максимальной силы через 5-30 мс.
Каждое отдельное мышечное волокно подчиняется закону «все или ничего», т. е. при силе раздражения выше порогового уровня происходит полное сокращение с максимальной для данного волокна силой, а ступенчатое повышение силы сокращения по мере увеличения силы раздражения невозможно. Поскольку смешанная мышца состоит из множества волокон с различным уровнем чувствительности к возбуждению, сокращение всей мышцы может быть ступенчатым в зависимости от силы раздражения, при этом при сильных раздражениях происходит активация глубжележащих мышечных волокон.
Механизм скольжения филаментов[править]
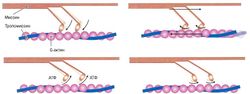 рис. 1. Схема образования поперечных связей — молекулярной основы сокращения саркомера
рис. 1. Схема образования поперечных связей — молекулярной основы сокращения саркомера Укорочение мышцы происходит за счет укорочения образующих ее саркомеров, которые, в свою очередь, укорачиваются за счет скольжения относительно друг друга актиновых и миозиновых филаментов (а не укорочения самих белков). Теория скольжения филаментов была предложена учеными Huxley и Hanson (Huxley, 1974; рис. 1). (В 1954 г. две группы исследователей — X. Хаксли с Дж. Хэнсон и А. Хаксли с Р. Нидергерке — сформулировали теорию, объясняющую мышечное сокращение скольжением нитей. Независимо друг от друга они обнаружили, что длина диска А оставалась постоянной в расслабленном и укороченном саркомере. Это позволило предположить, что есть два набора нитей — актиновые и миозиновые, причем одни входят в промежутки между другими, и при изменении длины саркомера эти нити каким-то образом скользят друг по другу. Сейчас эта гипотеза принята почти всеми.)
Актин и миозин — два сократительных белка, которые способны вступать в химическое взаимодействие, приводящее к изменению их взаимного расположения в мышечной клетке. При этом цепочка миозина прикрепляется к актиновой нити с помощью целого ряда особых «головок», каждая из которых сидит на длинной пружинистой «шее». Когда происходит сцепление между миозиновой головкой и актиновой нитью, конформация комплекса этих двух белков изменяется, миозиновые цепочки продвигаются между актиновыми нитями и мышца в целом укорачивается (сокращается). Однако, чтобы химическая связь между головкой миозина и активной нитью образовалась, необходимо подготовить этот процесс, поскольку в спокойном (расслабленном) состоянии мышцы активные зоны белка актина заняты другим белком — тропохмиозином, который не позволяет актину вступить во взаимодействие с миозином. Именно для того, чтобы убрать тропомиозиновый «чехол» с актиновой нити, требуется быстрое выливание ионов кальция из цистерн саркоплазматического ретикулума, что происходит в результате прохождения через мембрану мышечной клетки потенциала действия. Кальций изменяет конформацию молекулы тропомиозина, в результате чего активные зоны молекулы актина открываются для присоединения головок миозина. Само это присоединение осуществляется с помощью так называемых водородных мостиков, которые очень прочно связывают две белковые молекулы — актин и миозин — и способны в таком связанном виде находиться очень долго.
Для отсоединения миозиновой головки от актина необходимо затратить энергию аденозинтрифосфа-та (АТФ), при этом миозин выступает в роли АТФазы (фермента, расщепляющего АТФ). Расщепление АТФ на аденозиндифосфат (АДФ) и неорганический фосфат (Ф) высвобождает энергию, разрушает связь между актином и миозином и возвращает головку миозина в исходное положение. В дальнейшем между актином и миозином могут снова образовываться поперечные связи.
При отсутствии АТФ актин-миозиновые связи не разрушаются. Это и является причиной трупного окоченения (rigor mortis) после смерти, т. к. останавливается выработка АТФ в организме — АТФ предотвращает мышечную ригидность.
Даже при мышечных сокращениях без видимого укорочения (изометрические сокращения, см. выше) активируется цикл формирования поперечных связей, мышца потребляет АТФ и выделяет тепло. Головка миозина многократно присоединяется на одно и то же место связывания актина, и вся система миофиламентов остается неподвижной.
Внимание: Сократительные элементы мышц актин и миозин сами по себе не способны к укорочению. Мышечное укорочение является следствием взаимного скольжения миофиламентов относительно друг друга (механизм скольжения филаментов).
Как же образование поперечных связей (водородных мостиков) переходит в движение? Одиночный саркомер за один цикл укорачивается приблизительно на 5-10 нм, т.е. примерно на 1 % своей общей длины. За счет быстрого повторения цикла поперечных связей возможно укорочение на 0,4 мкм, или 20% своей длины. Поскольку каждая миофибрилла состоит из множества саркомеров и во всех них одновременно (но не синхронно) образуются поперечные связи, суммарно их работа приводит к видимому укорочению всей мышцы. Передача силы этого укорочения происходит через Z-линии миофибрилл, а также концы сухожилий, прикрепленных к костям, в результате чего и возникает движение в суставах, через которые мышцы реализуют перемещение в пространстве частей тела или продвижение всего тела.
Связь между длиной саркомера и силой мышечных сокращений[править]
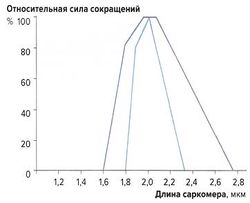 рис. 2. Зависимость силы сокращений от длины саркомера
рис. 2. Зависимость силы сокращений от длины саркомера Наибольшую силу сокращений мышечные волокна развивают при длине 2-2,2 мкм. При сильном растяжении или укорочении саркомеров сила сокращений снижается (рис. 2). Эту зависимость можно объяснить механизмом скольжения филаментов: при указанной длине саркомеров наложение миозиновых и актиновых волокон оптимально; при большем укорочении миофиламенты перекрываются слишком сильно, а при растяжении наложение миофиламентов недостаточно для развития достаточной силы сокращений.
Скорость укорочения мышечных волокон[править]
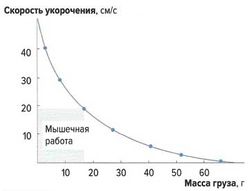 рис.3. Зависимость скорости укорочения от нагрузки
рис.3. Зависимость скорости укорочения от нагрузки Скорость укорочения мышцы зависит от нагрузки на эту мышцу (закон Хилла, рис. 3). Она максимальна без нагрузки, а при максимальной нагрузке практически равна нулю, что соответствует изометрическому сокращению, при котором мышца развивает силу, не изменяя своей длины.
Влияние растяжения на силу сокращений: кривая растяжения в покое[править]
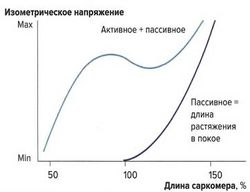 рис. 4. Влияние предварительного растяжения на силу сокращения мышцы. Предварительное растяжение повышает напряжение мышцы. Результирующая кривая, описывающая взаимоотношения длины мышцы и силы ее сокращения при воздействии активного и пассивного растяжения, демонстрирует более высокое изометрическое напряжение, чем в покое
рис. 4. Влияние предварительного растяжения на силу сокращения мышцы. Предварительное растяжение повышает напряжение мышцы. Результирующая кривая, описывающая взаимоотношения длины мышцы и силы ее сокращения при воздействии активного и пассивного растяжения, демонстрирует более высокое изометрическое напряжение, чем в покое Важным фактором, влияющим на силу сокращений, является величина растяжения мышцы. Тяга за конец мышцы и натяжение мышечных волокон называются пассивным растяжением. Мышца обладает эластическими свойствами, однако в отличие от стальной пружины зависимость напряжения от растяжения не линейна, а образует дугообразную кривую. С увеличением растяжения повышается и напряжение мышцы, но до определенного максимума. Кривая, описывающая эти взаимоотношения, называется кривой растяжения в покое.
Данный физиологический механизм объясняется эластическими элементами мышцы — эластичностью сарколеммы и соединительной ткани, располагающимися параллельно сократительным мышечным волокнам.
Также при растяжении изменяется и наложение друг на друга миофиламентов, однако это не оказывает влияния на кривую растяжения, т. к. в покое не образуются поперечные связи между актином и миозином. Предварительное растяжение (пассивное растяжение) суммируется с силой изометрических сокращений (активная сила сокращений).
- ↑ Harrison BC. et al. 2011. lib or not lib? Regulation of myosin heavy chain gene expression in mice and men. Skeletal Muscle. 1 (1): 5. doi: 10.1186/2044-5040-1-5.
- ↑ Andersen, J.L., et al. 1994. Myosin heavy chain isoforms in single fibres from m. vastus lateralis of sprinters: Influence of training. Acta Physiologica Scandinavica 151 (2): 135-42.
- ↑ 3,03,1 Andersen T.L, Aagaard P. 2000. Myosin heavy chain IIX overshoot in human skeletal muscle. Muscle Nerve. 23 (7): 1095-104.
- ↑ Andersen, L.L., et al. 2010. Early and late rate of force development: Differential adaptive responses to resistance training? Scandinavian Journal of Medicine and Science in Sports 20 (1): el62-69. doi:10.1111/j.l600-0838.2009.00933.x.
- ↑ Anderson, K., and Behm, D.G. 2004. Maintenance of EMG activity and loss of force output with instability. Journal of Strength and Conditioning Research 18:637-40.
- ↑ Aagaard, R, et al. 2011. Effects of resistance training on endurance capacity and muscle fiber composition in young top-level cyclists. Scandinavian Journal of Medicine and Science in Sports 21 (6): e298-307. doi:10.1111/j. 1600-0838.2010.01283.x.
- ↑ Henneman, E., Somjen, G., and Carpenter, D.O. 1965. Functional significance of cell size in spinal motoneurons./. Neurophysiol. 28:560-580.
- ↑ 8,08,1 Latash, M.L. 1998. Neurophysiological basis of movement. Champaign, IL: Human Kinetics.
- ↑ Brooks, G.A., Fahey, T.D., and White, T.P. 1996. Exercise physiology: Human bioenergetics and its applications. 2nd ed. Mountainview, CA: Mayfield.
- ↑ Schmidtbleicher, D. 1992. Training for power events. In Strength and power in sport, ed. P.V. Komi, 381-95. Oxford, UK: Blackwell Scientific.
- ↑ Wiemann, K., and Tidow, G. 1995. Relative activity of hip and knee extensors in sprinting—Implications for training. New Studies in Athletics 10 (1): 29-49.
sportwiki.to
Тренинг по типу мышечных волокон и их строение
Строение мышечных волокон
Каждая скелетная мышца состоит из пучков мышечных волокон, которые содержат множество сократительных нитей, миофибрилл.
У людей может быть разное количество волокон в мышцах. Максимальная сила увеличивается при большем числе волокон.

По морфологическим признакам и физиологическим свойствам мышечные волокна разделяют на быстрые и медленные. Их еще называют медленносокращающиеся и быстросокращающиеся. МС-волокна (ST) называют красными из-за миоглобина, придающим им красный мышечный пигмент. А БС-волокна (FT), или белые, обладают небольшим содержание миоглобина. БС-волокна могут развить силу в 10 раз большую, чем МС-волокна.
МС-волокнам свойственен аэробный окислительный обмен. В них присутствует много митохондрий, ферментов биологического окисления углеводов и жиров, миоглобина, который запасает кислород, у них наименьший диаметр, извилистая и широкая Z-линия. Они окружены множеством кровеносных капилляров. Соответственно, они идеальны для длительной работы.
На рисунке приведена организация митохондрий (множественные зубчатые черные линии).

У БС-волокон имеется много миофибрилл и большие запасы гликогена. В них мало миоглобина и капиллярная сеть плохо развита. Они хороши для короткой по времени (из-за утомления) интенсивной работы.
В свою очередь БС-волокна подразделяются на виды а и б, или FTO и FTG. FTO и FTG различаются по способу получения энергии. БС а-волокна («быстрые окислительно-гликолитические волокна») ресинтезируют АТФ анаэробным гликолитическим и аэробным путями. БС б-волокна получают энергию посредством гликолиза (глюкоза в анаэробных условиях распадается до лактата). Они хороши для короткой по времени мышечной деятельности интенсивного характера.
Характеристика этих волокон описана в таблице:


Набор МС- и БС-волокон индивидуален у каждого человека, и их количество нельзя изменить. Примерное соотношение: 40% медленных и 60% быстрых волокон у человека. Несмотря на то, что тренировкой не изменить соотношение БС и МС волокон, все же волокна приспосабливаются под воздействием физических нагрузок, например, изменяется их поперечное сечение, оснащение энергоносителями и митохондриями.
Интенсивность нагрузки
Последовательность включения различных мышечных волокон регулируется нервной системой и зависит от интенсивности нагрузки:
- 20-25% от максимальной силы- МС-волокна
- 25-40%- БС типа а (FTO)
- Более 40%- БС типа б (FTG)
Кстати, обычно после тренировок, в последующие дни, у всех нас болят мышцы, которые мы тренировали. О том, почему болят мышцы после тренировки, можно прочесть на нашем сайте в этом же разделе.
На графике продемонстрировано, как процент нагрузки от максимальной силы влияет на включение различных волокон в мышечную работу.

На нижеследующим графике изображен «эффект рампы» (Wilmore and Costill, 1999) на примере пловца. На нем видно, что все большее число волокон пловца подключается при увеличении скорости преодоления дистанции. FTa-волокна подключатся при большем усилии, когда увеличивается скорость. FTx (FTG) последними подключаются. Это происходит, когда скорость околомаксимальная.

Принцип величины (size principle)
Использование волокон при нагрузке зависит от размера мотонейрона и порога возбуждения. Эта зависимость называется «size principle» (Henneman, 1974). Малые мотонейроны МС-волокон вовлекаются первыми, так как их порог возбуждения низкий. Далее при увеличении нагрузки привлекаются более крупные мотонейроны. Самые большие мотонейроны с высоким порогом возбуждения- у БС б-волокон и поэтому они вовлекаются в работу последними. Малые мотонейроны иннервируют медленные двигательные единицы, которые развивают малую силу, устойчивы к утомлению и хороши для длительной работы. Крупные мотонейроны иннервируют быстрые двигательные единицы. Их сила велика, но они не устойчивы к длительной работе.
Таким образом, волокна вовлекаются в работу при большем усилии, которое прилагает спортсмен. Например, у спринтеров хорошо развиты мышцы ног, так как развиваемая ими скорость вынуждает быстрее сокращать мышцы и тем самым в работу скорее вовлекаются волокна с большим порогом возбуждения.
Допустим, у атлета одно единичное максимальное повторение для жима штанги лежа на скамье - 250 фунтов. Если он выжимает 125 фунтов, то будут вовлечены медленные двигательные единицы. Если жим будет с весом 225 на 6 повторений, то включатся и БС-волокна.
Есть ещё хороший пример с разгрузкой дивана (Jacob Wilson). Допустим, вы передвигаете диван в новый дом. Ваша цель - это перенести диван от грузовика до дома, и при этом преодолеть 10 ступенек. На первой ступеньке ноги начнут привлекать БС а-волокна. На второй/третьей - все больше БС а-волокон привлекается. Далее для поддержания бодрого темпа восхождения по ступенькам начнут привлекаться БС б-волокна. И на последней ступеньке должен наступить «отказ». Это применимо и к тренировочным сетам - в начале сета мышцы сокращаются не с максимальной частотой. Но в конце, с накоплением усталости, мышцы затрачивают больше силы, чтоб продолжать преодолевать нагрузку.
Но есть и подтверждения исключения из правила «size principle», когда мотонейроны БС-волокон вовлекаются в работу первее мотонейронов МС-волокон (Denier van der Gon, 1985; Grimby & Hannerz, 1977; Nardone, 1989; Smith, 1980; Ter Haar Romeny, 1982). Это происходит при выполнении эксцентрических упражнений.
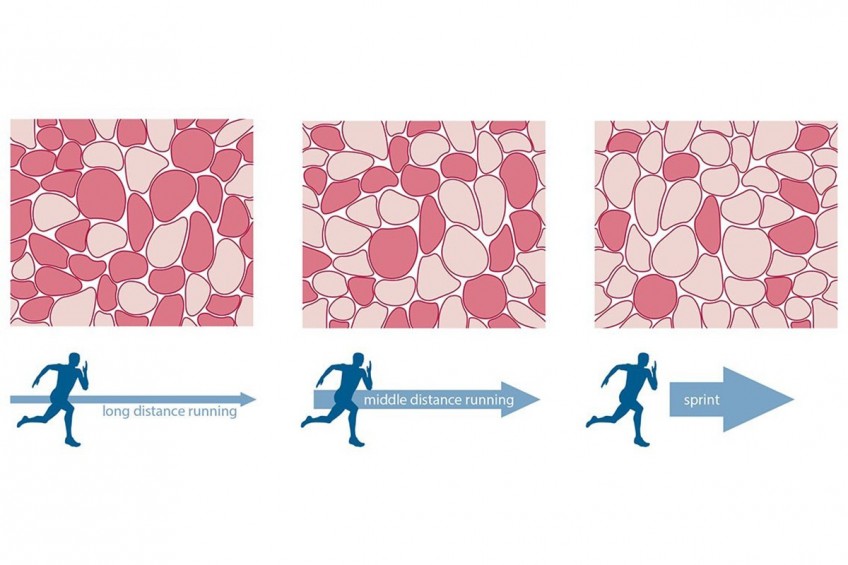
Уровень тренированности
Также, на привлечение волокон при физической активности влияет уровень тренированности. Например, у нетренированных людей включается примерно 60% волокон. А у силовиков - около 80-90%. Ниже приведены графики, отражающие вовлечение в работу волокон нетренированных (а) и тренированных (б) людей, 1 - МС-волокна, 2 - БС типа а-волокна, 3 - БС типа б, 4 - неиспользованные волокна.

Определение типов волокон
 Для прямого метод определения соотношения волокон у человека существует метод - биопсия. Но также существует и косвенный метод определения БС- и МС-волокон, основанный на связи между соотношением БС-волокон и мышечной силой (Coyle, 1979; Froese & Houston, 1985; Gerdle, 1988; Gregor, 1979; Suter, 1993). Сначала определяется максимальный вес в одном повторении (тот вес, с которым можно сделать одно повторение). Далее берется нагрузка в размере 80% от максимального веса единичного повторения и делается максимально возможное количество повторений. Если вышло меньше 7 повторений, то мышечная группа содержит более 50% БС-волокон. Если 12 повторений, то более 50% - это МС-волокна. Если 7-12 повторений, то, предполагается, что соотношение волокон равное.
Для прямого метод определения соотношения волокон у человека существует метод - биопсия. Но также существует и косвенный метод определения БС- и МС-волокон, основанный на связи между соотношением БС-волокон и мышечной силой (Coyle, 1979; Froese & Houston, 1985; Gerdle, 1988; Gregor, 1979; Suter, 1993). Сначала определяется максимальный вес в одном повторении (тот вес, с которым можно сделать одно повторение). Далее берется нагрузка в размере 80% от максимального веса единичного повторения и делается максимально возможное количество повторений. Если вышло меньше 7 повторений, то мышечная группа содержит более 50% БС-волокон. Если 12 повторений, то более 50% - это МС-волокна. Если 7-12 повторений, то, предполагается, что соотношение волокон равное.
Этот метод работает только для мышечных групп, так как работа с весами привлекает использование множества мышц. Для определения соотношения волокон в отдельной мышце требуется прямой метод биопсии.
Селективная гипертрофия, основанная на виде тренинга
Например, у атлета соотношение волокон 50/50. Но у БС-волокон площадь поперечного сечения больше, поэтому они могут занимать около 65% места в мышце. 35% приходится на МС-волокна. Поэтому программы на развитие силы не изменяют соотношение волокон, но изменяют поперечное сечение. Это происходит из-за того, что МС-волокна атрофируются (станут меньше), а БС-волокна гипертрофируются (станут больше). Также у атлета будет прибавка в массе, так как масса БС-волокон больше, чем у МС.
Если же спортсмен тренируется на выносливость, то БС-волокна атрофируются, а МС-волокна гипертрофируются, занимая больше места. Соотношение может измениться, например, с 65% БС-волокон и 35% МС-волокон на 50%/50% соответственно. При этом спортсмен потеряет мышечную массу, так как МС-волокна обладают меньшей массой.
Рост мышц (по Jacob Wilson)
В работе о мышечных волокнах, Jacob Wilson высказал некоторые рекомендации по поводу роста мышечной массы.
Тело приспосабливается к нагрузкам для выживания, а не для того, чтоб хорошо выглядеть на пляже. Дополнительная мышечная масса - это и дополнительная нагрузка для организма. Поэтому если возможно увеличить силу без гипертрофии, то это в первую очередь будет сделано. Чем больше мышечной массы, тем больше калорий требуется для ее поддержания, что не практично. А организм всегда поступает практично.
Автор приводит механизм адаптации к различным нагрузкам. Например, рассмотрим адаптацию к малым повторениям (1-5). Данная система хороша для развития силы. Нервная система должна реагировать моментально на такую нагрузку и привлекать как можно больше двигательных единиц. Поэтому адаптационные изменения будут происходить в нервной системе. Спортсмен научится работать с большим весом более эффективно, координируя все мышечные волокна. Поэтому пауэрлифтеры такие сильные. Чем больше двигательных единиц включается в работу, тем больший вес можно преодолеть. В данном диапазоне повторений не происходит желаемой мышечной гипертрофии для бодибилдера. Меньшее количество повторений не привлекает к работе и не утомляет все мышечные волокна.
Каждый тип волокон имеет определенный порог возбуждения. Для его подключения и роста требуется адекватная стимуляция. Значит, сет должен быть достаточно тяжел и длителен для достижения успехов в наборе массы. В таблице приведена зависимость количества повторений (с весом, позволяющим выполнить нужное количество повторений) и показателей роста волокон:
|
Кол-во повторений |
Показатель роста волокон |
|||
|
МС |
БС а |
БС б |
сила |
|
|
1-2 |
очень низкий |
низкий |
низкий |
отлично
|
|
3-5 |
очень низкий |
низкий |
хороший |
отлично |
|
6-8 |
очень низкий |
хороший |
отличный |
хороший |
|
9-12 |
низкий |
отличный |
очень хороший |
хороший |
|
13-15 |
неплохой |
очень хороший |
хороший |
выносливость |
|
16-25 |
очень хороший |
убывающий |
низкий |
выносливость |
|
25-50 |
отличный |
низкий |
очень низкий |
выносливость |
Среднее количество повторений позволяет задействовать большее количество волокон. Т.е. адаптация происходит не в направлении «чем больше двигательных единиц, тем лучше», а в постепенном привлечении волокон. К концу сета уже все волокна активизируются под нагрузкой. Также БС-волокна приобретают некоторые качества, присущие выносливым волокнам (повышается количество митохондрий).
В научной литературе описано, что аэробные, буфферные возможности и скорость удаление лактата в БС б-волокнах возрастает с уровнем тренированности, когда они начинают функционировать как БС а-волокна. Было произведена исследовательская работа, где в эксперименте принимала участие группа студентов (Anderson, 2005). Они тренировались с отягощениями 8 недель: жим ногами, гакк-приседания, сгибания/разгибания. Начинали они тренироваться на 10-12 повторений, но по мере увеличения веса отяжелений уменьшалось количество повторений (6-8).
Результаты биопсии представлены в таблице:
|
тип волокна |
до тренировок |
после |
разница |
|
МС |
60 |
60 |
0 |
|
БС а |
34 |
39 |
+5 |
|
БС б |
6 |
1 |
-5 |
Из этого следует, что БС а-волокна увеличиваются после тренировок с отягощениями. Тем самым увеличивается аэробные и анаэробные системы энергообеспечения. Что благоприятно сказывается на гипертрофии мышц.
По поводу МС волокон было установлено, что любая нагрузка (на выносливость, спринт) увеличивает их скорость сокращения и, следовательно, силу. Тренировки с отягощениями увеличивают их размер, что также сказывается на силе (MacDougall, 1986).
Любопытно еще одно открытие (Bonen, 2006). При тренировке МС-волокон происходит возрастание содержания вещества MCT1, транспортирующего лактат из крови и примыкающих БС-волокон в МС, где он метаболизируется митохондриями. Таким образом, ацидоз, провоцирующий усталость, наступает позднее в БС-волокнах.
Митохондрии
В них углеводы, жиры и протеины могут распадаться в присутствии кислорода, выделяя энергию. Они являются «топками» для сжигания жира.
Митохондрии увеличиваются при очень низкоинтенсивных или высокоинтенсивных нагрузках. На увеличение количества митохондрий влияют два параметра: увеличение кальция и уменьшение концентрации АТФ. Высокая концентрация кальция активизирует кальций-кальмодулин киназу, которая способствует росту митохондрий. Лучше всего такое увеличение митохондрий происходит при длительных упражнениях с малой скоростью.
Сокращение концентрации АТФ вызывает возрастание АМФ (аденозинмонофосфат), что активизирует АМФ-зависимую протеин-киназу, которая также запускает механизм увеличения количества митохондрий. Данная реакция происходит при высокоинтенсивной нагрузке.
Таким образом, низкоинтенсивные нагрузки хороши для улучшения аэробных способностей МС-волокон. В то время как высокоинтенсивные тренировки повышают аэробные способности БС-волокон.
БС с - волокна
Некоторые ученые не исключают возможности существования еще одного типа волокон: БС с, которое может посредством тренировок переходить в БС- а или МС-волокна (Wilmore, Costill, Kenney, 2008). Другие исследования также подтвердили наличие волокна-гибрида, которое сочетает характеристики МС и БС а-волокон (Brooks, 2005).
Но данная теория требует дальнейших исследований.
Выводы:
Тема мышечных волокон еще не достаточно хорошо изучена. Это подтверждает предположение о существовании БС с-волокон.
Знания о волокнах помогают определить специализацию спортсмена, принимая во внимание его генетические особенности.
Для мышечной гипертрофии важна нагрузка, которая достаточно тяжелая и длительная. Но важно помнить, что гипертрофия- это многофакторный процесс, зависящий от иммунного ответа на воспалительный процесс микротравмы, клеток-сателлитов, от факторов роста, гормонального фона, питания и пр.
Вовлечение волокон в работу зависит от интенсивности нагрузки, утомления и от усилия. «size principle» ошибочно отождествлять с «чем тяжелее, тем лучше», так как важным в привлечении волокон является усилие, которое испытывает спортсмен при выполнении упражнения. Если по окончании сета с любым количеством повторений наступает «отказ», то это значит, что БС-волокна были привлечены.
МС-волокнам свойственен аэробный окислительный обмен, а БС - анаэробный. Аэробный механизм энергообеспечения является основным при длительной работе умеренной мощности. При этом сокращается ограниченное число мышечных волокон. Аэробные упражнения способствуют сжиганию жира.
Анаэробный тренинг способствует развитию силы, наращиванию мышечной массы. Анаэробная система энергообеспечения включается при высокоинтенсивных и непродолжительных нагрузках. Программы на развитие силы не изменяют соотношение волокон, но изменяют поперечное сечение, причем у БС-волокон площадь поперечного сечения больше.
Автор: lena555
Используемая литература:
1. Muscle fibers, Jacob Wilson2. The size principle and a critical analysis of the unsubstantiated heavier-is-better recommendation for resistance training, Ralph N. Carpinelli3. The mystery of skeletal muscle hypertrophy, Len Kravitz, Richard Joshua Hernandez4. Muscle fiber types and training, Jason R. Karp5.Training Fast Twitch Muscle Fibers: Why and How, Ernest W. Maglischo6. Мышечная боль, Хабиров Ф.А.7. Биохимия мышечной деятельности, Волков Н.И., Несен Э.Н., Осипенко А.А., Корсун С.Н.8. Тренинг, питание, спортивная фармакология в бодибилдинге, Василенко А.
bituha.ru
Строение мышечного волокна — Мегаобучалка
Каждое мышечное волокно представляет собой гигантскую многоядерную клетку – симпласт, образованную в процессе эмбрионального развития организма путем слияния множества отдельных клеток – миобластов.
Строение мышечного волокна существенно отличается от строения других клеток. Важнейшие отличительные особенности - это размеры, форма, многоядерность, наличие сократительного аппарата. Строение мышечного волокна представлено на рис. 60.
Рис. 60. Важнейшие структурные элементы мышечного волокна
Остановимся на важнейших структурных элементах мышечного волокна.
Сарколемма.Снаружи мышечное волокно окружено оболочкой – сарколеммой, обладающей высокой прочностью и эластичностью. Эти свойства сарколеммы обеспечиваются присутствием в ней большого количества эластичных волокон белков коллагена и эластина, образующих густую сеть.
Сарколемма обладает избирательной проницаемостью, пропуская внутрь клетки преимущественно те вещества, для превращения которых там имеются условия – ферментные системы. В сарколемме присутствуют специальные транспортные системы, с помощью которых поддерживается, в частности, разница в концентрации ионов Na+, К+, Сl‾ внутри и снаружи мышечного волокна, что обеспечивает формирование на его поверхности мембранного потенциала.
К каждому мышечному волокну подходит окончание двигательного нерва. Место прикрепления нервного окончания к мышечному волокну называется нервно-мышечным синапсом..
Внутри мышечного волокна находятся многочисленные клеточные органеллы, важнейшими из которых являются ядра, митохондрии, рибосомы и др. Функции указанных органелл описаны в главе 2 (2.5.1). Пространство между органеллами заполнено внутриклеточной жидкостью – саркоплазмой. Среди структурных элементов мышечного волокна наибольший объем занимают сократительные нити– миофибриллы.
Миофибриллы. Миофибриллы представляют собой длинные тонкие нити, расположенные вдоль мышечного волокна. Количество миофибрилл в мышечных волокнах может колебаться в диапазоне от нескольких десятков до полутора и более тысяч. Под влиянием систематической мышечной тренировки, особенно скоростно-силовой направленности, количество миофибрилл может увеличиваться. Напротив, ограничение двигательной активности сопровождается уменьшением количества миофибрилл. Строение мышечных миофибрилл представлено на рис. 61 Рис. 61. Строение миофибрилл
При рассмотрении в оптический микроскоп видно, что миофибриллы имеют повторяющуюся поперечную исчерченность – темные и светлые полосы (диски). Темные диски (А-диски) в центральной части имеют более светлую полосу (Н-зону). Светлые диски (I-диски) в центре пересекаются узкой темной полосой (линией Z). Участок миофибрилл между двумя линиями Z получил название саркомера. Количество саркомеров в миофибрилле зависит от длины мышечного волокна и может достигать нескольких сотен. Длина саркомеров у разных людей может различаться.
Исследование срезов мышечных волокон в электронном микроскопе показали, что каждая миофибрилла состоит из большого числа параллельно расположенных толстых и тонких нитей (филаментов), которые характеризуются строгим взаимным распределением. Толстые нити находятся в зоне А-дисков. Они построены из белка миозина. Миозин является важнейшим сократительным белком, на его долю приходится около 55% от общего количества сократительных белков. Молекула миозина имеет длинную фибриллярную (удлиненную) часть и глобулярную (округлой формы) головку. Фибриллярная часть имеет двухспиральную полипептидную конфигурацию. Функция фибриллярной части молекулы миозина связана с формированием структуры толстой миозиновой нити.
Глобулярные головки миозиновых нитей имеют два активных центра, один из которых обладает АТФ-азной активностью (способностью расщеплять молекулы АТФ), другой – способностью связываться с активными центрами на актиновых нитях (актинсвязывающий центр). Головки молекул миозина располагаются на поверхности миозиновых нитей, образуя выпячивания (отростки). При этом они строго ориентированы в пространстве – располагаются шестью продольными рядами. Толстая миозиновая нить состоит как бы из двух частей, зеркально повторяющих друг друга. Если ее разрезать по- середине, то образуются два совершенно одинаковых фрагмента.
Молекулы миозина обладают способностью связывать ионы Са2+ и Мg2+. Ионы кальция являются кофактором фермента АТФ-азы (в его отсутствии фермент не активен). Ионы магния обеспечивают миозину способность связывать молекулы АТФ и АДФ.
В зоне светлых дисков (I-дисков) расположены тонкие нити, построенные из белков актина, тропомиозина и тропонина. Актин – второй в количественном отношении сократительный белок, составляющий основу актиновых нитей. Тропомиозин – структурный белок актиновых нитей, имеющий фибриллярную форму. Сдвоенные молекулы тропомиозина обвивают актиновые нити. Тропонин является регуляторным белком актиновых нитей. Он существует в трех формах, одна из которых блокирует взаимодействие актина с миозином. Другая форма способна связывать ионы кальция, благодаря чему изменяется конформация молекул первой формы тропонина и открывается центр взаимодействия актина с миозином. Третья форма тропонина обеспечивает крепление первых двух форм на актиновой нити. Кроме того, в составе тонких актиновых нитей имеется белок актинин. Он содержится в зоне линии Z, выполняющей роль своеобразной перегородки, и обеспечивает прикрепление к ней концов актиновых нитей.
К числу важнейших структурных элементов мышечного волокна относится саркоплазматический ретикулум. Саркоплазматический ретикулум - это внутриклеточная система взаимосвязанных пузырьков и канальцев (цистерн), пронизывающих клетку и особенно плотно концентрирующихся в зоне соприкосновения актиновых и миозиновых нитей.
Саркоплазматический (в клетках других органов и тканей – эндоплазматический) ретикулум имеется в каждой клетке организма человека. Но в мышечном волокне он выполняет несколько необычные по сравнению с другими клетками функции. Основная его роль в мышечном волокне заключается в регуляции содержания ионов кальция возле актиновых и миозиновых нитей. В состоянии расслабления ретикулум связывает ионы Са2+, их концентрация в саркоплазме составляет примерно 10-7 моль·литр-1. Под воздействием двигательного импульса ионы кальция освобождаются из ретикулума и их концентрация повышается до 10-5 моль·литр-1.
Способность саркоплазматического ретикулума связывать и высвобождать в цитоплазму ионы Са2+ связана с локализацией на его внутренней поверхности особых кальций связывающих белков. На поверхности ретикулума располагаются также рибосомы – особые внутриклеточные образования, в которых осуществляется синтез белков.
Мышечное волокно имеет также систему трубчатых выпячиваний сарколеммы (Т-систему), направленных внутрь мышечного волокна и располагающихся между миофибриллами и саркоплазматическим ретикулумом. Т-система обеспечивает быстрое распространение волны возбуждения от сарколеммы вглубь волокна.
В мышечном волокне содержатся и другие внутриклеточные органеллы: митохондрии, лизосомы. Функции этих структур мышечного волокна уже были описаны в главе «Общие закономерности обмена веществ».
Мышечное волокно содержит не одно, а несколько ядер, которые располагаются не в центральной части волокна, а по периметру, непосредственно под сарколеммой
Типы мышечных волокон
В скелетных мышцах выделяют несколько типов мышечных волокон, различающихся по своим двигательным характеристикам, соотношению различных химических и структурных компонентов, особенностям структурной организации. К основным типам мышечных волокон относятся медленносокращающиеся (МС) и быстросокращающиеся (БС). Медленносокращающиеся волокна в связи с более высоким содержанием в них миоглобина называют еще красными (или тип I). Быстросокращающиеся, для которых характерно более низкое содержание миоглобина, называют белыми (или тип II). Следует сразу оговориться, что различить эти два типа волокон по цвету практически невозможно. Цвет у одних и у других красный.
Бытрые и медленные волокна более чем в два раза различаются по максимальной скорости сокращения. Так, время одиночного сокращения МС достигает 110 мс, а БС – 50 мс. Кроме того, БС более чем в два раза превышают МС по своим силовым характеристикам.
Существенно различаются разные типы волокон по уровню развития различных механизмов преобразования энергии. МС волокна имеют хорошо развитый механизм аэробного ресинтеза АТФ, что обеспечивается большим количеством митохондрий и высоким содержанием ферментов аэробного биологического окисления, а также большими запасами субстратов аэробного окисления: гликогена, жиров. В МС волокнах содержится больше белка миоглобина, благодаря которому они имеют больший запас кислорода и более благоприятные условия для перехода кислорода из крови внутрь волокна.
В БС волокнах значительно больше миофибрилл, выше АТФ-азная активность, больше концентрация ионов кальция.
Внутри БС волокон различают два подтипа: БСа и БСб. Эти два подтипа отличаются, главным образом, разным уровнем развития важнейших механизмов преобразования энергии. У волокон БСа более хорошо развит анаэробный гликолиз и несколько слабее, чем у МС волокон - аэробный путь ресинтеза АТФ. Они являются ведущими при выполнении упражнений т.н. субмаксимальной мощности, продолжительность которых колеблется в диапазоне от 30 сек до 2-3 мин, при условии выполнения работы с максимальной для данной продолжительности интенсивностью.
У волокон БСб, наряду с анаэробным гликолизом хорошо развит креатинфосфатный механизм ресинтеза АТФ. Они подключаются при выполнении упражнений максимальной и околомаксимальной интенсивности: бег на 100 м, упражнения с большими отягощениями и т.п.
Сказанное не означает, что упражнения указанной интенсивности, выполняются исключительно одним типом мышечных волокон. Речь идет о степени их вовлечения в работу, что, безусловно, определяется центральной нервной системой. Выполнение упражнений, мощность которых не превышает 20-25% от максимально возможной для данного индивидуума обеспечивается только «красными» мышечными волокнами. При работе, интенсивность которой находится в диапазоне 25-40% от максимальной, к ее выполнению подключаются волокна БСа. Если интенсивность упражнения превышает 40% от максимальной, в работу вовлекаются волокна БСб.
При повышении интенсивности упражнения в пределах каждой зоны мощности увеличивается участие в ее обеспечении волокон всех типов, но в наибольшей степени тех, которые подключаются к работе в данном диапазоне мощности.
Различные типы мышечных волокон различаются и условиями иннервации. Мотонейроны, иннервирующие БС мышечные волокна, более толстые, они имеют более разветвленную сеть нервных окончаний (ветвей аксонов), благодаря чему иннервируют значительно большее количество мышечных волокон (от300 до 500). У БС волокон больше зона прилегания нервного окончания к мышечному волокну, что создает более благоприятные условия для иннервации и возникновения потенциала действия.
megaobuchalka.ru