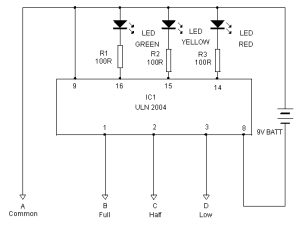
Васкуляризация красных и белых мышечных волокон. Среди каких мышц выделяют белые и красные мышцы
Как правильно качать мышцы белые и красные (Таблица).
Когда я бываю в тренажерном зале, то замечаю, что многие «пирожки» вообще не понимают, что они делают и для чего. Чтобы ваши тренировки были осознанными и вы понимали какие мышцы вы развиваете, а главное как это нужно делать я подготовил статью и таблицу «Как правильно качать мышцы белые и красные».
На теле человека более 600 различных мышц. 50% мышц сосредоточено в нижних конечностях, 30% — в верхних конечностях и 20% приходится на мышцы головы и туловища. У женщин масса мышц составляет 30-35% от массы тела, а у мужчин 40-45%, у спортсменов 45-55%.
Различают «быстрые» мышцы (ГМВ), «красные» мышцы (ОМВ).
Гликолитические мышечные волокна (ГМВ) хорошо подходят для взрывной силы, скорости, спринтерского бега. Окислительные мышечные волокна (ОМВ) наоборот, больше приспособлены для выносливости, длительного бега.
Тренируются каждый тип мышечных волокон по-разному (см. Таблицу). В динамическом режиме развивают ГМВ, а в статодинамическом ОМВ. Если в тренировке ГМВ используют полную амплитуду движения (присел-встал), то при тренировке ОМВ используют небольшую амплитуду движения под напряжением, порядка 15-20% (присели – чуть привстали, снова чуть присели). Подробнее смотрите в таблице «Как правильно качать мышцы белые и красные?»
Между подходами необходимо отдыхать для восстановления ГМВ 5-10 минут, а ОМВ 3-7 минут. Здесь подразумевается активный отдых. Это не сидеть на стуле, а покрутить педали, походить подвигаться, чтобы выгнать из мышцы ионы водорода. В это время можно делать подходы на другую группу мышц. Например, делали ноги, а пока перерыв, то можно делать на руки или пресс.
3 повтора на группу мышц – поддерживающая тренировка. Если хотите, чтобы мышцы развивались, тогда необходимо делать от 4 до 9 повторов на каждую группу мышц, которую хотите развивать.
Развивающую тренировку достаточно делать 1 раз в две недели, чтобы мышцы могли полноценно развиваться. Но если вам очень хочется, то можно 1 раз в неделю, но не чаще на конкретную группу мышц.
Как правильно качать мышцы (белые и красные).
Похожее
maximbuvalin.ru
Красные и белые мышцы | Здоровье, быт, увлечения, отношения
 Мышечные волокна, в зависимости от утомляемости, гистохимической окраски и сократительных свойств, делят на две группы – белые и красные мышцы.
Мышечные волокна, в зависимости от утомляемости, гистохимической окраски и сократительных свойств, делят на две группы – белые и красные мышцы.
Медленными волокнами небольшого диаметра являются красные мышечные волокна. Чтобы получать энергию они используют окисление жирных кислот и углеводов. Также эти волокна называют: SТ-волокна, волокна первого типа, медленносокращающиеся и медленные мышечные волокна.
Эти волокна имеют красную гистохимическую окраску, которая обусловлена большим содержанием в них миоглобина, доставляющего вглубь мышечного волокна и в кровь кислород.
В красных волокнах содержится большое количество митохондрий. В них для получения энергии и происходит процесс окисления.
Сила сокращений медленных мышечных волокон сравнительно невелика, также им достаточно для энергии аэробного метаболизма. Для таких волокон подходит продолжительная и не интенсивная работа (аэробика, ходьба или легкий бег, небольшие дистанции в плавании), движения, которые не требуют поддержания позы и значительных усилий. Красные мышцы отличаются превосходной выносливостью, они начинают работать при нагрузке от максимальной силы в пределах двадцати-двадцати пяти процентов.
Медленные волокна не подходят для больших дистанций в плавании, подъема тяжелого веса, потому что для таких нагрузок требуется быстрое получение и расход энергии.
Диаметр белых мышечных волокон больше, чем диаметр красных. Они являются быстрыми, а для получения энергии используют гликолиз. Также их называют: FТ-волокна, волокна второго т
ипа, быстросокращающиеся мышечные волокна, быстрые.
Волокна выглядят белее, потому что в них меньше миоглобина. В белых мышцах вырабатывается большое количество энергии, так как активность фермента АТФазы высока. Скорость расхода энергии в этих волокнах очень высока, поэтому и скорость восстановления АТФ должна быть высокой, что и обеспечивает процесс гликолиза, который проходит в саркоплазме мышечных волокон. Белые мышцы быстро устают, так как способны при гликолизе накапливать молочную кислоту (лактат). В итоге мышца останавливает свою работу.
В белых волокнах содержится меньшее, по сравнению с красными волокнами, количество митохондрий, но большее количество гликогена и миофибрилл. Также в них находится, необходимый в начале высокоинтенсивной работы, креатинфосфат.
Быстрые волокна подходят для кратковременных, но мощных и быстрых действий. FT-волокна развивают большую в десять раз силу, чем красные волокна, а также сокращаются в два раза быстрее.
Быстрые белые волокна бывают двух типов:
-Быстросокращающиеся гликолитические волокна. Для получения энергии они используют процесс гликолиза. Скорость сокращения и сила быстрых гликолитических волокон максимальная. Их роль в обеспечении бегунам и пловцам максимальной скорости – первостепенна.
-Быстросокращающиеся окислительно-гликолитические волокна. Они являются промежуточными или переходными быстрыми волокнами. Такие волокна имеют мощную анаэробную систему энергообразования. Однако они способны выполнять достаточно интенсивную аэробную работу.
goldstarinfo.ru
Васкуляризация красных и белых мышечных волокон
Со времени опубликования работ Spalteholz (1888) и Krogh (1919) сложилось мнение, что микроциркуляторное русло скелетных мышц имеет достаточно строгую пространственную упорядоченность и сравнительно простое однообразное строение. Так, модель, предложенная Hammersen (1968), предполагает кристаллоподобное строение скелетной мышцы. Между тем известно, что любая скелетная мышца по составу мышечных волокон представляет собой гетерогенную структуру, при этом белые (или быстрые, фазные) и красные (медленные, тонические) мышечные волокна имеют различную степень васкуляризации.
Продолжение ниже ⇓В 70-х годах прошлого столетия Ranvier (1874), наблюдая скелетные мышцы кролика, обратил внимание на то, что мышечные волокна имеют разную скорость сокращения. Еще до открытия быстрых и медленных мышц мышечные волокна дифференцировались по цвету на красные и белые. Разная окраска мышечных волокон связывалась с неодинаковым содержанием миоглобина в мышечной ткани.
По ориентировочным данным, белые мышечные волокна (БМВ) у человека составляют около 80-85% от общей мышечной массы. Кровоток в них равен 40-60 мл/100 г/мин. При тяжелой мышечной работе он увеличивается до 15-16 л/мин. Возможно, что у спортсменов он примерно на 30% выше (Folkow, Neil, 1971).
Красные мышечные волокна (КМВ), которым свойственны длительные сокращения, используются преимущественно при статических напряжениях и во время длительной равномерной активности (скажем, при поддержании позы). Имеются указания (Hilton, 1966), что объем сосудистого русла в тонических мышцах больше, чем в фазных; кровоток в красных мышцах в покое составляет около 100-150 мл/100 г/мин, что примерно в 2-2,5 раза выше, чем в белых мышцах.
Различия диаметров мышечных волокон удовлетворительно коррелируют с особенностями их гистологического строения. Между тем поиск морфологических особенностей васкуляризации красных и белых мышц осложняется значительными техническими трудностями, связанными с идентификацией состава мышечных волокон в изучаемых мышцах.
Разработка методов гистохимической окраски мышцы показала, что белые волокна отличаются высокой активностью фосфорилазы, альдолазы, пируваткиназы, лактатдегидрогеназы и аглицерофос-фатдегидрогеназы, а в красных волокнах хорошо выявляются цито-хромоксидазы и ферменты, окисляющие пировиноградную, янтарную и изолимонную кислоты (Ogata, 1960; George, Talesara, 1961; Beat-ty et al,, 1963; Romanul, 1964, 1965).
При изучении спектра ферментов в мышцах Stein, Padycula (1962), Ogata, Mari (1964), Hemman, Olson (1965) в качестве основных гистохимических методов для дифференцировки мышечных волокон использовали окраску на гликоген, АТФ и неспецифическую эстеразу. По степени активности ферментов Nachmias, Padycula (1958) выделили у крыс три типа мышечных волокон А, В, С (белые, промежуточные и красные). Romanul (1964), изучая икроножную и камбаловидную мышцы крысы и кролика, сначала выделил 8 типов волокон, а позднее (Romanul, 1965) объединил их в три группы. В икроножной мышце им были найдены волокна всех трех типов (А, В и С), а в камбаловидной - волокна типа В и С.
Исследуя камбаловидную, икроножную и четырехглавую мышцы крыс, В. В. Португалов с соавт. (1968, 1971) отмстили их неоднородность по составу волокон. Так, в состав камбаловидной мышцы входят два типа волокон - красные и промежуточные; в состав икроножной и четырехглавой мышц - красные, промежуточные и белые волокна с диаметром 27, 37 и 44 мкм (соответственно).
Красные мышечные волокна, как отмечает Denny-Brown (1929), имеют тенденцию группироваться в обособленные глубоколежащие головки. По данным В. И. Дерибас (1967, 1969), в четырехглавой мышце бедра и передней большеберцовой мышце у белой крысы глубоколежащие мышечные пучки состоят на 77% из красных мышечных волокон и на 23% из белых; поверхностно расположенные мышечные пучки - на 26% из красных и па 74% из белых мышечных волокон. Процентное содержание в скелетной мышце того или иного типа мышечных волокон определяет, по-видимому, ее способность производить сокращения фазного или тонического типа.
Для аэробного метаболизма красных мышечных волокон необходим усиленный приток кислорода. Поэтому различия в потреблении кислорода мышцами зависят от состава мышечных волокон. Приведенные. В. И. Дерибас (1967, 1969) абсолютные величины «дыхания» только красных и только белых мышечных волокон в четырехглавой мышце бедра и в передней большеберцовой мышце белой крысы свидетельствуют о том, что «дыхание» красных мышечных волокон во много раз интенсивнее, чем белых. Так, потребление кислорода красными мышечными волокнами составляло 24 мл/100 г/мин, а белыми - 0,6 мл/100 г/мин.
Поскольку красные мышцы требуют в покое большего притока кислорода, кровоснабжение их должно превосходить кровоснабжение белых мышц. По данным Reis et al (1967), объем кровотока на 100 г мышечной ткани в красных мышечных волокнах составил 28-31 мл/мин, а в белых - 10-12 мл/мин. Аналогичные данные приводят Hilton et al. (1970), которые установили, что в покое кровоток в камбаловидной (преимущественно красной) мышце у кошки составляет 51,9 мл/100 г/мин, а в икроножной (преимущественно белой) - 13,9 мл/100 г/мин, т. е. примерно в 4 раза меньше.
Несомненный интерес для характеристики васкуляризации красных и белых мышечных волокон имеет расчет плотности функционирующих капилляров. Ж. Т. Искакова (1977, 1978, 1979), проводя исследование в нашей лаборатории, точно морфометрически определила плотность общего числа капилляров и числа функционирующих капилляров в четырехглавой мышце бедра белой крысы и кошки с учетом зон преимущественной локализации красных и белых мышечных волокон. Для выявления количественных характеристик капиллярного русла она использовала два принципиально различных метода: один из них - тотальная инъекция сосудов раствором туши - позволил выявить все капилляры в мышце, другой - окраска эритроцитов бензидином - лишь те капилляры, которые функционируют в мышце в тот или иной момент времени, предшествующий фиксации материала. Подсчет общего числа капилляров и числа функционирующих капилляров производился на поперечных срезах мышцы (прямой и латеральной головок четырехглавой мышцы бедра) с учетом зон преимущественного расположения красных и белых мышечных волокон.
Чтобы определить эти зоны, предварительно была проведена гистохимическая окраска поперечных срезов мышцы па выявление сукцинатдегидрогеназы. В результате было установлено, что в центральной зоне прямой головки мышцы преимущественно содержатся красные мышечные волокна, а в латеральной головке и наружных слоях прямой головки - белые мышечные волокна.
Как оказалось, наиболее информативными параметрами являются не абсолютные показатели плотности капилляров, которые подвержены значительным индивидуальным и видовым колебаниям, а относительные показатели, полученные в перерасчете на число мышечных волокон, определяемых на единицу площади поперечного среза мышцы. Расчет среднего диаметра мышечного волокна показал, что он хорошо коррелирует с размерами мышцы. Чем больше вес мышцы, тем больше средний диаметр мышечных волокон.
Применение инъекционной методики, выявляющей все капилляры в мышце, и методики окраски поперечных срезов мышцы бензидином, выявляющей лишь функционирующие капилляры, позволило, во-первых, установить различия в васкуляризации красных и белых мышечных волокон, а во-вторых, сопоставить число функционирующих и нефункционирующих капилляров в скелетной мышце и оценить резервные возможности капиллярного русла.
Расчет плотности капилляров в четырехглавой мышце бедра позволил определить, что общее число капилляров в зоне преимущественного расположения КМВ составляет 2184 на 1 мм2 (78±7,8 на поле зрения) поверхности поперечного сечения, а в зоне преимущественного расположения БМВ -1092 на 1 мм2 (39±5,6 на поле зрения). Плотность функционирующих капилляров в покоящейся мышце оказалась значительно ниже и составляет в зоне КМВ 896 на 1мм2 (32±1,9 на поле зрения), в зоне БМВ -336 на 1 мм2 (12±1,2 на поле зрения). Значит, в покоящейся четырехглавой мышце бедра у белой крысы функционирует лишь часть капилляров в зоне расположения КВМ и часть капилляров в зоне расположения БМВ. Следовательно, резервные возможности капиллярного русла в скелетной мышце достаточно велики, что и обеспечивает сравнительно высокую функциональную лабильность и приспособляемость капиллярного кровотока при изменении рабочего состояния мышцы. Поэтому структурные параметры микроциркуляторного русла не могут служить фактором, лимитирующим изменчивость капиллярного кровотока.
Наиболее информативным представляется поверхностный показатель (ПП), который характеризует отношение поверхности капилляров к поверхности обслуживаемых ими мышечных волокон. Он рассчитывается по формуле, которую приводит К. А. Шошенко (1975): где NK-число капилляров, NMB-число мышечных волокон, радиус капилляра (в наших расчетах принимался равным 2,5 мкм), радиус мышечного волокна. Если различия мышечных волокон и их плотность зависят от размеров самой мышцы и веса животного, то, как показали исследования Ж. Т. Исскаковой (1979), ПП существенно зависит только от особенностей васкуляризации мышечных волокон. Зависимость ПП от веса тела животного, рассчитанная по методу наименьших квадратов. Из рисунка следует, что в этом показателе индивидуальные различия васкуляризации отдельных мышц снивелированы; показаны принципиальные различия в кровоснабжении красных и белых мышечных волокон.
© Авторы и рецензенты: редакционный коллектив оздоровительного портала "На здоровье!". Все права защищены.
www.nazdor.ru
Красные мышцы по массе уступают белым. Их количество у большинства рыб составляет несколько процентов от массы белой мускулатуры. И лишь у активных пловцов, находящихся в постоянном движении - тунцов, ставриды, макрели, сельди -доля красной мускулатуры достигает 20 %.Красная мускулатура находится над белой мускулатурой, непосредственно под кожей, в основном в латеральной области хвостового стебля и туловища рыбы.Лишь у видов, использующих для движения грудные плавники, красная мускулатура локализована в непосредственной близости от плавников и жаберных крышек. У активных пловцов (например, тунца) красные мышцы пронизывают всю толщу белых мышц и достигают позвоночного столба (рис. 5.8).Темный цвет мускулатуре придают железосодержащие белки. Мышечные волокна содержат большое количество миоглобина и цитохромов. Кроме того, темная мускулатура имеет обильную васкуляризацию. В состоянии покоя кровоток в красных мышцах в 3 раза превышает таковой в белых мышцах. При возбуждении приток крови к темным мышцам многократно возрастает.
§23. ГЛАДКИЕ МЫШЦЫГладкие мышцы образуют стенки внутренних трубчатых органов. Микроструктура этого типа мышц не так однородна, как микроструктура скелетной мускулатуры. У них нет поперечной исчерченности, так как нет упорядочение расположенных толстых и тонких протофибрилл. Во многих висцеральных мышцах толстых протофибрилл вообще нет. Однако при химическом анализе мыши в них обязательно обнаруживается белок миозин. Считают, что в последнем случае сокращение мышечного волокна происходит за счет полимеризации молекул миозина.В составе некоторых гладких мышц обнаружены очень толстые Темные протофибриллы сложного строения. Их основу составляет белок тропомиозин, а на поверхности находится миозин.Помимо специфики фибриллярного аппарата гладкие мышцы отличаются от поперечнополосатых еще и слаборазвитой тубулярной системой. Так, у гладких мышц нет Т-трубочек. Саркоплазматическая система слабо развита (в некоторых мышцах вообще отсутствует) и представлена своеобразными пузырьками, Протофибриллярный аппарат имеет непосредственные зоны соприкосновения с сарколеммой.Волокна гладких мышц могут быть как длинными, так и короткими. У рыб и те и другие образуют контакты друг с другом. Не все волокна в гладкой мышце имеют контакты с нервным окончанием. Поэтому двигательная единица гладких мыши имеет специфическое строение. Она включает нервное окончание вегетативного нерва, концевую пластинку и группу из 5-10 мышечных клеток, которые имеют контакты между собой. Естественно, что такая единица реагирует на нервную стимуляцию очень медленно.Гладкие мышцы способны к спонтанному возбуждению. Большинство висцеральных органов имеет интрамуральные нервные сплетения, которые обеспечивают высокую степень автономности трубчатых органов.Гладкие мышцы в отличие от скелетных не чувствительны к электрической стимуляции и очень чувствительны к механическому раздражению (например, растяжению). Отличаются гладкие мышцы и своей высокой реактивностью по отношению к химическим раздражителям (ацетилхолин, норадреналин. окситоцин, соли калия, кальция, магния, иногда желчь, соляная кислота), что не свойственно поперечнополосатым мышцам.Сокращения гладких мышц носят характер медленных и продолжительных, что и требуется для обеспечения перемешивания и продвижения химуса по пищеварительной трубке, выделения желчи, мочи, извержения половых продуктов при икрометании и т. д.
§24. МЕХАНИЗМ МЫШЕЧНОГО СОКРАЩЕНИЯСобытия, развивающиеся на микроскопическом (субклеточном) уровне при возбуждении мышцы, выглядят следующим образом. Потенциал действия, возникающий в результате нервной или иной стимуляции на мембране мышечного волокна, в скелетных мышцах по Т-каналам достигает мембран цитоплазматического ретикулума. Мембрана последнего высвобождает ионизированный кальций, который, в свою очередь, активизирует процесс взаимодействия актиновых и миозиновых протофибрилл.В гладких мышцах, лишенных тубулярной системы, высвобождение Са++ происходит непосредственно в самой сарколемме. А так как сарколемма напрямую контактирует с актомиозиновым комплексом гладких мышц, то активизация сокращения гладких мышц также происходит под влиянием ионизированного кальция.Интимный механизм взаимодействия актиновых и миозиновых структур до конца не расшифрован. Однако ясно, что в присутствии АТФ в поперечнополосатых мышцах при выбросе Са++ начинается скольжение тонких и толстых протофибрилл относительно друг друга (рис. 5.12). Считается, что непосредственной причиной этого движения протофибрилл является активизация актомиозиновых мостиков, которые либо изменяет угол наклона, либо подвергаются торсионному скручиванию (рис. 5.12). На поверхности воды летучая рыба достаточно продолжительно работает хвостом, развивая большую тягу, позволяющую ей преодолеть силу притяжения. Скорость полета этих мелких рыб превышает скорость движения преследователей (тунцы, меч-рыба), а пролетаемые ими расстояния достигают нескольких сотен метров.Другие виды рыб, например пальцекрыл, могут не только парить, но и выполнять сложные маневры в воздухе. Пальцекрыл поднимается к поверхности воды и скользит по ней со скоростью 18м/с. Такую высокую скорость рыба приобретает благодаря зигзагообразным движениям хвостового плавника с гипертрофированной нижней лопастью (рис. 5.15).Сильный удар хвоста поднимает рыбу в воздух на высоту 5-7 м. Пальцекрыл пролетает в воздухе до 200м, используя при этом и воздушные потоки. Рыба способна при необходимости изменить направление полета за счет движений хвостового плавника. У нее также отмечены колебательные движения грудных плавников.Скорость полета пальцекрыла сопоставима со скоростью движения современных морских судов и нередко достигает 60- 70 км/ч,Интересен механизм движений, приводящий в "боевое" положение колючки плавников таких рыб, как ерши, окуни, колюшки, сомы. Острые и крепкие колючки плавников этих рыб защищают их от хищников.В случае опасности плавники расправляются, а колючки поднимаются перпендикулярно телу. При этом несколько увеличиваются границы тела рыбы. Проглотить или схватить такую рыбу может не каждый хищник, так как надавливание на колючки не приводит к их складыванию. Следовательно, хищник при захвате неизбежно травмирует ротовую полость. Вероятнее всего, в следующий раз хищник откажется от нападения на такую рыбу.
§25. СКОРОСТЬ ДВИЖЕНИЯСкорость плавания рыбы в воде пропорциональна частоте и амплитуде колебаний тела и хвоста и укладывается в рамки определенных математических уравнений. Например, для карася, ельца и форели предельная скорость плаванияv=(1/4)[L(3f-4)],где L - длина тела; f- частота колебаний тела (хвоста).Несмотря на то что вода является более плотной средой по сравнению с воздухом, рыбам удается достичь очень высоких скоростей (км/ч):Рыба-меч 130Голубой марлин 90 Голубой тунец 80Терпуг 70Голубая акула 60Атлантический лосось 40Естественно, что для обеспечения таких скоростей природа наделила рыб рядом морфологических, гидродинамических и метаболических приспособлений.Поскольку предельные абсолютные скорости рыб зависят от их чиненных размеров, специалисты предлагают использовать для сопоставления скоростных возможностей рыб относительный показатель - коэффициент скорости:Этот коэффициент характеризует скорость рыбы, равную числу ее корпусов (длин) в секунду. Тогда все виды рыб можно классифицировать, как минимум, шестью категориями (табл. 5.4). Понятно, что рыбы с одинаковыми максимальными скоростями движения, но с различной длиной тела согласно этой классификации могут относиться к разным категориям. |
 Рис. 5.8. Топография красной мускулатуры на поперечном разрезе тела тунцаФункционально красная мускулатура также выделяется. Она обеспечивает мед. ленную, но продолжительную работу. Красные мышцы меньше утомляются и быстрее восстанавливаются при интенсивных нагрузках. Именно в такого рода мышечной деятельности испытывают повышенную потребность активные пелагические рыбы - тунцы и макрели, покрывающие за сутки расстояния в сотни и тысячи километров.В качестве источника энергии красные мышцы используют жирные кислоты и кетоновые тела, что возможно лишь при наличии кислорода. В красных мышцах содержится мало гликогена и много ферментов, необходимых для аэробных реакций. Даже при предельном напряжении в красных мышцах активно протекают аэробные процессы. Поэтому так велика потребность красных мышц в кислороде и так развита в них сеть капиллярных сосудов.Мышечные волокна красной мускулатуры иннервируются медленными нервными волокнами, т. е. волокнами малого диаметра. Это частично объясняет причины большого латентного периода красных мышц. Однако на каждом мышечном волокне имеется несколько концевых пластинок, гарантирующих точность выполнения команды из центральной нервной системы.Светлые мышцы у рыб составляют основную массу активной части опорно-двигательного аппарата. Они отличаются более коротким латентным периодом (по сравнению с темными мышцами), большой скоростью сокращения и расслабления. Как для светлых, так и для красных мышц характерны тетанические сокращения, что определено природой электрического импульса, поступающего по аксону мотонейрона, - спайк с частотой 30- 50 Гц. Иннервация светлых мышц осуществляется за счет быстрых (миелинизированных) нервных волокон большого диаметра. Каждое мышечное волокно у костистых рыб имеет несколько нервно-мышечных синапсов.Кровоснабжение светлых мышц не такое интенсивное, как красных. В качестве источника энергии светлые мышцы используют глюкозу. Тот факт, что в возбужденных светлых мышцах всегда присутствует молочная кислота, свидетельствует о приоритете гликолиза в процессе мышечного сокращения.Приверженность светлых мышц к анаэробному использованию энергии глюкозы подчеркивает и высокая активность ферментов анаэробного цикла в светлых мышцах рыб. Известно, что гликолиз - неэф-фективный путь использования энергии по сравнению с окислительным фосфорилированием. В данном случае природа приносит в жертву энергетическую экономию для получения выигрыша в скорости мышечного сокращения.Клетка мышечной ткани (мышечное волокно) имеет специфическое строение. Снаружи она покрыта особой мембранной оболочкой - сарколеммой, а ее протоплазма (саркоплазма) наряду с традиционными цитозольными структурами включает в себя особые органеллы - миофибриллы,Миофибрилла является структурным элементом мышечного волокна, обеспечивающим укорочение (сокращение) (рис. 5.9).При световом микроскопировании миофибрилл видна их поперечная исчерченность из-за разницы в светопреломлении входящих в их состав белков - актина и миозина. В состав миофибрилл входят толстые и тонкие протофибриллы. Тонкие актиновые протофибриллы образуют так называемые изотропные диски (I-диски). В средней части изотропного диска расположена Z-пластина.Более толстые миозиновые протофибриллы и заходящие своими концами актиновые нити образуют анизотропные диски (А-диски). В средней части анизотропных дисков тонких нитей нет, поэтому эту зону выделяют в самостоятельную Н-зону. В области взаимного перекрытия в пределах А-диска актиновые и миозиновые нити соединены при помощи особых мостиков. Кроме того, актиновые нити закреплены на поперечных Z-пластинах (рис. 5.10).
Рис. 5.8. Топография красной мускулатуры на поперечном разрезе тела тунцаФункционально красная мускулатура также выделяется. Она обеспечивает мед. ленную, но продолжительную работу. Красные мышцы меньше утомляются и быстрее восстанавливаются при интенсивных нагрузках. Именно в такого рода мышечной деятельности испытывают повышенную потребность активные пелагические рыбы - тунцы и макрели, покрывающие за сутки расстояния в сотни и тысячи километров.В качестве источника энергии красные мышцы используют жирные кислоты и кетоновые тела, что возможно лишь при наличии кислорода. В красных мышцах содержится мало гликогена и много ферментов, необходимых для аэробных реакций. Даже при предельном напряжении в красных мышцах активно протекают аэробные процессы. Поэтому так велика потребность красных мышц в кислороде и так развита в них сеть капиллярных сосудов.Мышечные волокна красной мускулатуры иннервируются медленными нервными волокнами, т. е. волокнами малого диаметра. Это частично объясняет причины большого латентного периода красных мышц. Однако на каждом мышечном волокне имеется несколько концевых пластинок, гарантирующих точность выполнения команды из центральной нервной системы.Светлые мышцы у рыб составляют основную массу активной части опорно-двигательного аппарата. Они отличаются более коротким латентным периодом (по сравнению с темными мышцами), большой скоростью сокращения и расслабления. Как для светлых, так и для красных мышц характерны тетанические сокращения, что определено природой электрического импульса, поступающего по аксону мотонейрона, - спайк с частотой 30- 50 Гц. Иннервация светлых мышц осуществляется за счет быстрых (миелинизированных) нервных волокон большого диаметра. Каждое мышечное волокно у костистых рыб имеет несколько нервно-мышечных синапсов.Кровоснабжение светлых мышц не такое интенсивное, как красных. В качестве источника энергии светлые мышцы используют глюкозу. Тот факт, что в возбужденных светлых мышцах всегда присутствует молочная кислота, свидетельствует о приоритете гликолиза в процессе мышечного сокращения.Приверженность светлых мышц к анаэробному использованию энергии глюкозы подчеркивает и высокая активность ферментов анаэробного цикла в светлых мышцах рыб. Известно, что гликолиз - неэф-фективный путь использования энергии по сравнению с окислительным фосфорилированием. В данном случае природа приносит в жертву энергетическую экономию для получения выигрыша в скорости мышечного сокращения.Клетка мышечной ткани (мышечное волокно) имеет специфическое строение. Снаружи она покрыта особой мембранной оболочкой - сарколеммой, а ее протоплазма (саркоплазма) наряду с традиционными цитозольными структурами включает в себя особые органеллы - миофибриллы,Миофибрилла является структурным элементом мышечного волокна, обеспечивающим укорочение (сокращение) (рис. 5.9).При световом микроскопировании миофибрилл видна их поперечная исчерченность из-за разницы в светопреломлении входящих в их состав белков - актина и миозина. В состав миофибрилл входят толстые и тонкие протофибриллы. Тонкие актиновые протофибриллы образуют так называемые изотропные диски (I-диски). В средней части изотропного диска расположена Z-пластина.Более толстые миозиновые протофибриллы и заходящие своими концами актиновые нити образуют анизотропные диски (А-диски). В средней части анизотропных дисков тонких нитей нет, поэтому эту зону выделяют в самостоятельную Н-зону. В области взаимного перекрытия в пределах А-диска актиновые и миозиновые нити соединены при помощи особых мостиков. Кроме того, актиновые нити закреплены на поперечных Z-пластинах (рис. 5.10).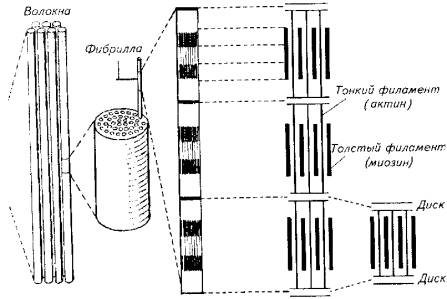 Рис. 5.9. Микроструктура скелетной мышцыИсследования позволили выявить в составе тонких актиновых нитей и элементы другого белка - миозина (в виде тропомиозина).Участок миофибриллы между двумя Z-пластинами называют саркомером.Количество и соотношение толстых и тонких протофибрилл колеблется не только у разных видов рыб, но и в разных мышцах у одной особи. Быстрые мышцы имеют большее количество нитей и меньше саркоплазмы. В медленных мышцах больше саркоплазмы.Помимо нитевидных структур - миофибрилл и протофибрилл, в составе мышечного волокна скелетных мышц выделяется хорошо развитая тубулярная система - сеть каналов и трубочек. Подсчитано, что общая поверхность мембран тубулярной системы в 10 раз превышает площадь наружной мембраны мышечного волокна. Сарколемма образует разветвленные выпячивания, проникающие в область Z-пластин. Их называют Т-канальцами. К Т-канальцам прилегает другая тубулярная система, которая относится к саркоплазматическому ретикулуму. Она образует сложную ячеистую сеть вокруг саркомера (рис. 5.11).Эта мембранная сеть обладает очень высокой ферментативной активностью. Она же выступает в роли депо ионизированного кальция, без которого невозможно мышечное сокращение.В медленных мышцах объем Т-канальцев значительно меньше по сравнению с их объемом в быстрых мышцах.Потенциал действия, возникающий на сарколемме при возбуждении мышцы, передается по Т-канальцам, а затем по каналам саркоплазматического ретикулума достигает непосредственно протофибриллярного аппарата.
Рис. 5.9. Микроструктура скелетной мышцыИсследования позволили выявить в составе тонких актиновых нитей и элементы другого белка - миозина (в виде тропомиозина).Участок миофибриллы между двумя Z-пластинами называют саркомером.Количество и соотношение толстых и тонких протофибрилл колеблется не только у разных видов рыб, но и в разных мышцах у одной особи. Быстрые мышцы имеют большее количество нитей и меньше саркоплазмы. В медленных мышцах больше саркоплазмы.Помимо нитевидных структур - миофибрилл и протофибрилл, в составе мышечного волокна скелетных мышц выделяется хорошо развитая тубулярная система - сеть каналов и трубочек. Подсчитано, что общая поверхность мембран тубулярной системы в 10 раз превышает площадь наружной мембраны мышечного волокна. Сарколемма образует разветвленные выпячивания, проникающие в область Z-пластин. Их называют Т-канальцами. К Т-канальцам прилегает другая тубулярная система, которая относится к саркоплазматическому ретикулуму. Она образует сложную ячеистую сеть вокруг саркомера (рис. 5.11).Эта мембранная сеть обладает очень высокой ферментативной активностью. Она же выступает в роли депо ионизированного кальция, без которого невозможно мышечное сокращение.В медленных мышцах объем Т-канальцев значительно меньше по сравнению с их объемом в быстрых мышцах.Потенциал действия, возникающий на сарколемме при возбуждении мышцы, передается по Т-канальцам, а затем по каналам саркоплазматического ретикулума достигает непосредственно протофибриллярного аппарата.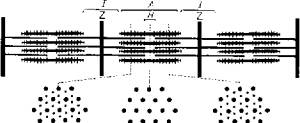 Рис. 5.10. Взаимное расположение актиновых и миозиновых протофибрилл
Рис. 5.10. Взаимное расположение актиновых и миозиновых протофибрилл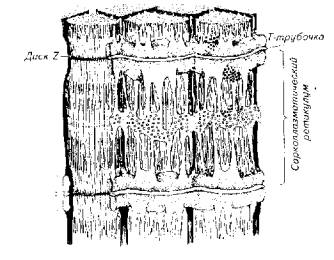 Рис. 5.11. Схема, иллюстрирующая расположение фибриллярных элементов и тубулярной сети мышечного волокнаВ целом скорость сокращения мышцы зависит от длины саркомера, количества протофибрилл и характера их расположения в пределах саркомера, количества митохондрий и энергетического материала, наличия и количества миоглобина в саркоплазме.
Рис. 5.11. Схема, иллюстрирующая расположение фибриллярных элементов и тубулярной сети мышечного волокнаВ целом скорость сокращения мышцы зависит от длины саркомера, количества протофибрилл и характера их расположения в пределах саркомера, количества митохондрий и энергетического материала, наличия и количества миоглобина в саркоплазме.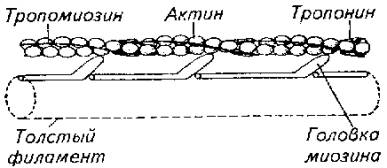 Рис. 5.12. Модель взаимодействия актиновой и миозиновой нитейУстановлена АТФ-азная зависимость конформационных изменений актомиозиновых мостиков. В отсутствие АТФ эти изменения невозможны, т. е. процесс скольжения протофибрилл не происходит.Следует подчеркнуть исключительно важную роль ионов кальция в этом процессе, который не только влияет на конформационные изменения актомиозина, но и является активатором АТФ-азы. Более того, процесс расслабления мышцы зависит от Са++. Актиновые и миозиновые нити не возвращаются в исходное положение до тех пор, пока не произойдет обратное поглощение ионов кальция мембраной цитоплазматического ретикулума или мембраной сарколеммы (в гладких мышцах), причем это обратное движение кальция зависит от активности АТФ-азы соответствующего мембранного комплекса. Поэтому не только сокращение, но и расслабление мышцы протекает с затратами энергии.Эксперименты показали, что АТФ-азную активность проявляет сам белок миозин. Он же при наличии АТФ способен соединяться с актином. Причем АТФ-азная часть миозина активируется актином. Однако белок тропонин в составе актиновой нити блокирует эту активность. И только выброс Са++ снимает тормозящее действие тропонина. Поэтому тропонин-тропомиозиновую систему рассматривают как предохранительный механизм взаимодействия актиновых и миозиновых нитей в состоянии физиологического покоя.Механизм укорочения гладкомышечного волокна менее понятен. Распространена точка зрения, согласно которой укорочение гладкомышечного волокна есть результат конформационного изменения актомиозинового комплекса. Белок актомиозин выделен из гладких мышц многих животных, включая рыб. По своим свойствам он близок к миозину млекопитающих. Как актиновая, так и миозиновая молекула имеют фибриллярную спиралевидную часть. Активизация актомиозина приводит к изменению конформации молекулы белка с соответствующим изменением его пространственного внутрицеллюлярного положения. Это и приводит к общему укорочению мышечного волокна, так как актомиозиновые структуры морфологически связаны с сарколеммой гладкомышечного волокна.Нервная регуляция гладкомышечной активности у рыб довольно специфична и заслуживает специального обсуждения. Влияние нервной системы на гладкие мышцы может быть как возбуждающим, так и тормозящим. Характер нервной стимуляции гладких мышц существенно зависит от гормонального статуса рыб и функционального состояния самого висцерального органа (например, степени наполнения желудка, мочеточников, стадии созревания половых клеток в ястыке и их наличия в матке или яйцеводе и т. д.).Гладкая мускулатура иннервируется симпатическим и парасимпатическим отделами нервной системы. Кроме того, многие висцеральные органы (например, желудочно-кишечный тракт крупные сосуды) обладают собственной интрамуральной нервной системой в виде нервных сплетений разной степени сложности.Как правило, симпатические нервные волокна в качестве медиаторов выделяют норадреналин, а парасимпатические волокна - соответственно ацетилхолин. Однако исследования показали, что в составе парасимпатической системы есть адренергические, а в составе симпатической системы -холинергические волокна. Поэтому у рыб симпатическая и парасимпатическая нервные системы являются смешанными по функциональным признакам. У ацетилхолина имеется два типа рецепторов - мускариновые и никотиновые. Для норадреналина описано также два функциональных типа рецепторов, называемых а- и р-рецепторами.Стимуляция вагуса как представителя парасимпатической части нервной системы у рыб вызывает различные эффекты не только по отношению к различным физиологическим системам, но и в пределах одной системы. Так, у форели и других желудочных рыб вагус тормозит работу желудка. Возбуждающему эффекту желудок подвергается при стимуляции чревного нерва.Далее желудка (кишечник) влияние симпатической и парасимпатической систем изменяется на прямо противоположное. Адренергические волокна и адреналин оказывают тормозящее действие, а холинергические волокна и ацетилхолин стимулируют перистальтику кишечника. Интересно и то, что ацетилхолин возбуждает все отделы кишечника у рыб. Адреналин же возбуждает только желудок и оказывает тормозящее действие на весь кишечник.У некоторых рыб пищеварительная трубка содержит элементы поперечнополосатой мускулатуры. Например, у линя кишка покрыта своеобразным чехлом подобных мышц. Поэтому его кишечник одинаково отвечает как на электрическую, так и на гуморальную стимуляцию.Кроме плавания и висцеральных функций мышцы обеспечивают и некоторые довольно специфичные движения рыб. Так, таиландский ходячий сом в составе грудных плавников имеет шипы-ходули, которыми он пользуется для переползания по суше из одного водоема в другой. Наличие таких ходуль и приспособленность к дыханию атмосферным воздухом (имеет хорошо развитую систему наджаберных полостей) позволяют этой рыбе преодолевать в период засухи довольно большие расстояния в поискахПравда, при движении по суше (точнее сказать грязи, так как рыба для передвижения все-таки выбирает увлажненные места) сом использует не только грудные плавники-ходули, но и при вытянутом теле, извиваясь подобно змее, облегчает работу мышц плавников. Эта особенность таиландского сома позволила ему освоить большие территории на юге США, куда он случайно попал из Азии.Морской петух тригла (рис. 5.13) также ползает по дну, но при этом опирается сразу на три луча грудных плавников (отсюда и его название "тригла"). Интересно, что эти ходильные лучи у петуха обособлены от других и способны сгибаться. Кроме движения по дну видоизмененные лучи грудных плавников обеспечивают этой рыбе тактильную рецепцию.В других случаях рыбы используют грудные плавники для планирования в воздухе. Так, летучие рыбы при испуге за счет броскового движения туловищных мышц, мышц хвостового стебля и интенсивной работы нижней лопастью хвостового плавника выскакивают из воды и пролетают в воздухе расстояния, по-зволяющие им избавиться от преследователей (рис. 5.14).
Рис. 5.12. Модель взаимодействия актиновой и миозиновой нитейУстановлена АТФ-азная зависимость конформационных изменений актомиозиновых мостиков. В отсутствие АТФ эти изменения невозможны, т. е. процесс скольжения протофибрилл не происходит.Следует подчеркнуть исключительно важную роль ионов кальция в этом процессе, который не только влияет на конформационные изменения актомиозина, но и является активатором АТФ-азы. Более того, процесс расслабления мышцы зависит от Са++. Актиновые и миозиновые нити не возвращаются в исходное положение до тех пор, пока не произойдет обратное поглощение ионов кальция мембраной цитоплазматического ретикулума или мембраной сарколеммы (в гладких мышцах), причем это обратное движение кальция зависит от активности АТФ-азы соответствующего мембранного комплекса. Поэтому не только сокращение, но и расслабление мышцы протекает с затратами энергии.Эксперименты показали, что АТФ-азную активность проявляет сам белок миозин. Он же при наличии АТФ способен соединяться с актином. Причем АТФ-азная часть миозина активируется актином. Однако белок тропонин в составе актиновой нити блокирует эту активность. И только выброс Са++ снимает тормозящее действие тропонина. Поэтому тропонин-тропомиозиновую систему рассматривают как предохранительный механизм взаимодействия актиновых и миозиновых нитей в состоянии физиологического покоя.Механизм укорочения гладкомышечного волокна менее понятен. Распространена точка зрения, согласно которой укорочение гладкомышечного волокна есть результат конформационного изменения актомиозинового комплекса. Белок актомиозин выделен из гладких мышц многих животных, включая рыб. По своим свойствам он близок к миозину млекопитающих. Как актиновая, так и миозиновая молекула имеют фибриллярную спиралевидную часть. Активизация актомиозина приводит к изменению конформации молекулы белка с соответствующим изменением его пространственного внутрицеллюлярного положения. Это и приводит к общему укорочению мышечного волокна, так как актомиозиновые структуры морфологически связаны с сарколеммой гладкомышечного волокна.Нервная регуляция гладкомышечной активности у рыб довольно специфична и заслуживает специального обсуждения. Влияние нервной системы на гладкие мышцы может быть как возбуждающим, так и тормозящим. Характер нервной стимуляции гладких мышц существенно зависит от гормонального статуса рыб и функционального состояния самого висцерального органа (например, степени наполнения желудка, мочеточников, стадии созревания половых клеток в ястыке и их наличия в матке или яйцеводе и т. д.).Гладкая мускулатура иннервируется симпатическим и парасимпатическим отделами нервной системы. Кроме того, многие висцеральные органы (например, желудочно-кишечный тракт крупные сосуды) обладают собственной интрамуральной нервной системой в виде нервных сплетений разной степени сложности.Как правило, симпатические нервные волокна в качестве медиаторов выделяют норадреналин, а парасимпатические волокна - соответственно ацетилхолин. Однако исследования показали, что в составе парасимпатической системы есть адренергические, а в составе симпатической системы -холинергические волокна. Поэтому у рыб симпатическая и парасимпатическая нервные системы являются смешанными по функциональным признакам. У ацетилхолина имеется два типа рецепторов - мускариновые и никотиновые. Для норадреналина описано также два функциональных типа рецепторов, называемых а- и р-рецепторами.Стимуляция вагуса как представителя парасимпатической части нервной системы у рыб вызывает различные эффекты не только по отношению к различным физиологическим системам, но и в пределах одной системы. Так, у форели и других желудочных рыб вагус тормозит работу желудка. Возбуждающему эффекту желудок подвергается при стимуляции чревного нерва.Далее желудка (кишечник) влияние симпатической и парасимпатической систем изменяется на прямо противоположное. Адренергические волокна и адреналин оказывают тормозящее действие, а холинергические волокна и ацетилхолин стимулируют перистальтику кишечника. Интересно и то, что ацетилхолин возбуждает все отделы кишечника у рыб. Адреналин же возбуждает только желудок и оказывает тормозящее действие на весь кишечник.У некоторых рыб пищеварительная трубка содержит элементы поперечнополосатой мускулатуры. Например, у линя кишка покрыта своеобразным чехлом подобных мышц. Поэтому его кишечник одинаково отвечает как на электрическую, так и на гуморальную стимуляцию.Кроме плавания и висцеральных функций мышцы обеспечивают и некоторые довольно специфичные движения рыб. Так, таиландский ходячий сом в составе грудных плавников имеет шипы-ходули, которыми он пользуется для переползания по суше из одного водоема в другой. Наличие таких ходуль и приспособленность к дыханию атмосферным воздухом (имеет хорошо развитую систему наджаберных полостей) позволяют этой рыбе преодолевать в период засухи довольно большие расстояния в поискахПравда, при движении по суше (точнее сказать грязи, так как рыба для передвижения все-таки выбирает увлажненные места) сом использует не только грудные плавники-ходули, но и при вытянутом теле, извиваясь подобно змее, облегчает работу мышц плавников. Эта особенность таиландского сома позволила ему освоить большие территории на юге США, куда он случайно попал из Азии.Морской петух тригла (рис. 5.13) также ползает по дну, но при этом опирается сразу на три луча грудных плавников (отсюда и его название "тригла"). Интересно, что эти ходильные лучи у петуха обособлены от других и способны сгибаться. Кроме движения по дну видоизмененные лучи грудных плавников обеспечивают этой рыбе тактильную рецепцию.В других случаях рыбы используют грудные плавники для планирования в воздухе. Так, летучие рыбы при испуге за счет броскового движения туловищных мышц, мышц хвостового стебля и интенсивной работы нижней лопастью хвостового плавника выскакивают из воды и пролетают в воздухе расстояния, по-зволяющие им избавиться от преследователей (рис. 5.14).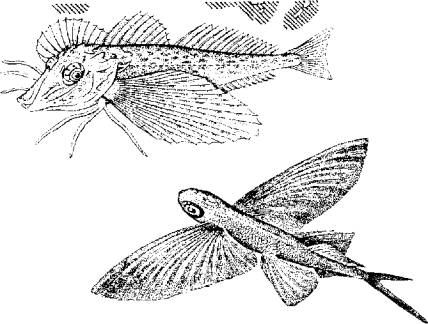 Рис. 5.13. Морской петух использует лучи грудных плавников для "хождения" по дну. Рис. 5.14. Полет пальцекрыла
Рис. 5.13. Морской петух использует лучи грудных плавников для "хождения" по дну. Рис. 5.14. Полет пальцекрыла Рис. 5.15. След на поверхности воды, оставляемый летучей рыбой при взлетеКроме того, расправленные плавники и торчащие колючки ложно увеличивают размеры рыбы, что также повышает ее шансы на выживание.Важно отметить, что колюшкам и сомам не приходится прибегать к мышечной силе, так как колючки в "боевом" положенийудерживаются не мышцами, а при помощи особого механизма. (Рис. 5.16.)B механизме фиксации колючки заложен принцип замка, который используется и другими животными (коленный сустав лошади, суставы нижних конечностей вороны и др.). Чем сильнее хищник давит на колючку, тем прочнее колючка фиксируется в замке, т е. прочность замка возрастает за счет использования энергии хищника, а не потенциальной жертвы.Опорой для колючки и ее замка служит особое образование из нескольких костей и мышц, формирующих шарнир (см. рис.5.16,///)Шарнир запирается при помощи V-образной косточки, которая имеет прочные связки с колючкой и скелетом. При возбуждении рыбы мышцы 5 и 6 сокращаются, колючка поднимается, а кость 3 блокирует шарнир. В таком положении давление на колючку делает фиксацию шарнира только жестче.Для опускания колючки необходимо, чтобы расслабились мышцы 5, 6 и 7, но сократилась мышца 8. На удержание колючки в поднятом состоянии расходуется минимум мышечных усилий, а для приведения в горизонтальное положение требуется сокращение всего одной мышцы 8.Рыбы-брызгуны используют мышечный аппарат глотки, рта и жабр для выплевывания струи воды (рис. 5.17).
Рис. 5.15. След на поверхности воды, оставляемый летучей рыбой при взлетеКроме того, расправленные плавники и торчащие колючки ложно увеличивают размеры рыбы, что также повышает ее шансы на выживание.Важно отметить, что колюшкам и сомам не приходится прибегать к мышечной силе, так как колючки в "боевом" положенийудерживаются не мышцами, а при помощи особого механизма. (Рис. 5.16.)B механизме фиксации колючки заложен принцип замка, который используется и другими животными (коленный сустав лошади, суставы нижних конечностей вороны и др.). Чем сильнее хищник давит на колючку, тем прочнее колючка фиксируется в замке, т е. прочность замка возрастает за счет использования энергии хищника, а не потенциальной жертвы.Опорой для колючки и ее замка служит особое образование из нескольких костей и мышц, формирующих шарнир (см. рис.5.16,///)Шарнир запирается при помощи V-образной косточки, которая имеет прочные связки с колючкой и скелетом. При возбуждении рыбы мышцы 5 и 6 сокращаются, колючка поднимается, а кость 3 блокирует шарнир. В таком положении давление на колючку делает фиксацию шарнира только жестче.Для опускания колючки необходимо, чтобы расслабились мышцы 5, 6 и 7, но сократилась мышца 8. На удержание колючки в поднятом состоянии расходуется минимум мышечных усилий, а для приведения в горизонтальное положение требуется сокращение всего одной мышцы 8.Рыбы-брызгуны используют мышечный аппарат глотки, рта и жабр для выплевывания струи воды (рис. 5.17).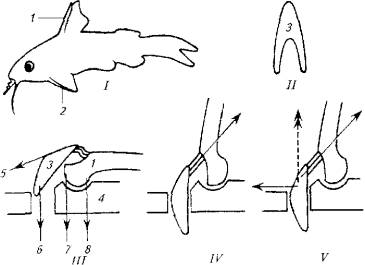 Рис. 5.16. Схема фиксации плавниковых колючек:I- общий вия рыбы; III- IV- разные положения сочленения костей; 1 - колючка спинного плавника; 2- колючка грудного плавника; 3- опорный скелет; 4, II-V-образная косточка; 5-8- места прикрепления мышц
Рис. 5.16. Схема фиксации плавниковых колючек:I- общий вия рыбы; III- IV- разные положения сочленения костей; 1 - колючка спинного плавника; 2- колючка грудного плавника; 3- опорный скелет; 4, II-V-образная косточка; 5-8- места прикрепления мышц Рис. 5.17. Рыба-брызгун: охота на насекомых при помощи струи водыРыба-брызгун достигла большого совершенства в прицельной стрельбе по насекомым, При длине собственного тела не более 20 см рыба выбрасывает мощную струю воды, которая сбивает насекомых на расстоянии до 1,5 м от поверхности воды. Впечатляет и высокая результативность такой охоты. Промахи у этой рыбы - крайне редкое явление.Все это разнообразие движений, наблюдаемое в классе рыб, обеспечивает сложная система опорно-двигательного аппарата и гладкомышечных образований, объединенных в единое целое при помощи нервной системы. Движения рыбы, в какой бы форме они ни проявлялись, осуществляют оперативное реагирование организма на изменения среды обитания с целью избежания опасности или адаптации к новым условиям.
Рис. 5.17. Рыба-брызгун: охота на насекомых при помощи струи водыРыба-брызгун достигла большого совершенства в прицельной стрельбе по насекомым, При длине собственного тела не более 20 см рыба выбрасывает мощную струю воды, которая сбивает насекомых на расстоянии до 1,5 м от поверхности воды. Впечатляет и высокая результативность такой охоты. Промахи у этой рыбы - крайне редкое явление.Все это разнообразие движений, наблюдаемое в классе рыб, обеспечивает сложная система опорно-двигательного аппарата и гладкомышечных образований, объединенных в единое целое при помощи нервной системы. Движения рыбы, в какой бы форме они ни проявлялись, осуществляют оперативное реагирование организма на изменения среды обитания с целью избежания опасности или адаптации к новым условиям.stydopedia.ru
Белые мышечные волокна | Анаэробная нагрузка | Увеличение мышечной массы
 У человека мышцы сформированы из двух типов мышечных волокон: белые (быстрые) и красные (медленные) волокна. Сегодня мы будем говорить о белых волокнах.
У человека мышцы сформированы из двух типов мышечных волокон: белые (быстрые) и красные (медленные) волокна. Сегодня мы будем говорить о белых волокнах.
Что из себя представляют белые мышечные волокна и для чего они нужны?
Думаю, небольшое количество людей задается данным вопросом. И раз вы на этом сайте, значит, какой-то интерес у вас всё-таки присутствует.
Итак, чтобы белые волокна имели возможность сокращаться, ими используется бескислородный способ получения энергии. Другими словами обмен веществ в белых мышечных волокнах протекает без участия кислорода. Белые волокна предназначены для осуществления быстрых, резких и взрывных усилий. Но при этом они имеют свойство быстро уставать. Зато максимальная скорость и сила достигается именно за счет быстрых волокон.
В свою очередь белые волокна делятся на два вида:
- Промежуточный вид (переходный). Этот вид сочетает в себе волокна для аэробных нагрузок (бег, плавание) и анаэробных нагрузок (короткие дистанции, тяжелая атлетика). Они начинают работать в тот момент, когда одновременно требуется сила и выносливость.
- И собственно сами быстрые волокна для силовых и скоростных сокращений мышц. Они также отвечают за увеличение мышечной массы. Все, кто посещает зал с целью увеличить силу и мышцы, работают с помощью этих мышечных волокон.
 Почему мышечный объем проще увеличить с быстрыми волокнами, потому что их диаметр гораздо больше диаметра красных медленных волокон. Соответственно, если у вас больше белых волокон, то и больше возможностей для роста мышц. Но это не значит, что нужно ограничиваться одними штангами. Красные волокна также требуют внимания. Применяйте аэробные нагрузки для их развития.
Почему мышечный объем проще увеличить с быстрыми волокнами, потому что их диаметр гораздо больше диаметра красных медленных волокон. Соответственно, если у вас больше белых волокон, то и больше возможностей для роста мышц. Но это не значит, что нужно ограничиваться одними штангами. Красные волокна также требуют внимания. Применяйте аэробные нагрузки для их развития.
В среднем, количество белых волокон у человека немного больше, чем красных, но всё индивидуально и соотношение может быть разным. Явное отличие в соотношение волокон можно наблюдать у спортсменов спринтеров или штангистов. У них количество белых (быстрых) волокон более 75%. Также у тех, у кого преобладают белые волокна, больше шансов добиться успехов в большом силовом спорте. В частности, в бодибилдинге. Не унывайте, если ваша физиология не позволяет похвастаться наличием белых мышечных волокон. На формирование стройного и здорового тела это никак не повлияет.
Рубрика: Бодибилдинг, Фитнес и здоровье |bodykeeper.ru
| АрхеологияАрхитектураАстрономияАудитБиологияБотаникаБухгалтерский учётВойное делоГенетикаГеографияГеологияДизайнИскусствоИсторияКиноКулинарияКультураЛитератураМатематикаМедицинаМеталлургияМифологияМузыкаПсихологияРелигияСпортСтроительствоТехникаТранспортТуризмУсадьбаФизикаФотографияХимияЭкологияЭлектричествоЭлектроникаЭнергетика |
Красные мышцы по массе уступают белым. Их количество у большинства рыб составляет несколько процентов от массы белой мускулатуры. И лишь у активных пловцов, находящихся в постоянном движении - тунцов, ставриды, макрели, сельди -доля красной мускулатуры достигает 20 %.Красная мускулатура находится над белой мускулатурой, непосредственно под кожей, в основном в латеральной области хвостового стебля и туловища рыбы.Лишь у видов, использующих для движения грудные плавники, красная мускулатура локализована в непосредственной близости от плавников и жаберных крышек. У активных пловцов (например, тунца) красные мышцы пронизывают всю толщу белых мышц и достигают позвоночного столба (рис. 5.8).Темный цвет мускулатуре придают железосодержащие белки. Мышечные волокна содержат большое количество миоглобина и цитохромов. Кроме того, темная мускулатура имеет обильную васкуляризацию. В состоянии покоя кровоток в красных мышцах в 3 раза превышает таковой в белых мышцах. При возбуждении приток крови к темным мышцам многократно возрастает.
§23. ГЛАДКИЕ МЫШЦЫГладкие мышцы образуют стенки внутренних трубчатых органов. Микроструктура этого типа мышц не так однородна, как микроструктура скелетной мускулатуры. У них нет поперечной исчерченности, так как нет упорядочение расположенных толстых и тонких протофибрилл. Во многих висцеральных мышцах толстых протофибрилл вообще нет. Однако при химическом анализе мыши в них обязательно обнаруживается белок миозин. Считают, что в последнем случае сокращение мышечного волокна происходит за счет полимеризации молекул миозина.В составе некоторых гладких мышц обнаружены очень толстые Темные протофибриллы сложного строения. Их основу составляет белок тропомиозин, а на поверхности находится миозин.Помимо специфики фибриллярного аппарата гладкие мышцы отличаются от поперечнополосатых еще и слаборазвитой тубулярной системой. Так, у гладких мышц нет Т-трубочек. Саркоплазматическая система слабо развита (в некоторых мышцах вообще отсутствует) и представлена своеобразными пузырьками, Протофибриллярный аппарат имеет непосредственные зоны соприкосновения с сарколеммой.Волокна гладких мышц могут быть как длинными, так и короткими. У рыб и те и другие образуют контакты друг с другом. Не все волокна в гладкой мышце имеют контакты с нервным окончанием. Поэтому двигательная единица гладких мыши имеет специфическое строение. Она включает нервное окончание вегетативного нерва, концевую пластинку и группу из 5-10 мышечных клеток, которые имеют контакты между собой. Естественно, что такая единица реагирует на нервную стимуляцию очень медленно.Гладкие мышцы способны к спонтанному возбуждению. Большинство висцеральных органов имеет интрамуральные нервные сплетения, которые обеспечивают высокую степень автономности трубчатых органов.Гладкие мышцы в отличие от скелетных не чувствительны к электрической стимуляции и очень чувствительны к механическому раздражению (например, растяжению). Отличаются гладкие мышцы и своей высокой реактивностью по отношению к химическим раздражителям (ацетилхолин, норадреналин. окситоцин, соли калия, кальция, магния, иногда желчь, соляная кислота), что не свойственно поперечнополосатым мышцам.Сокращения гладких мышц носят характер медленных и продолжительных, что и требуется для обеспечения перемешивания и продвижения химуса по пищеварительной трубке, выделения желчи, мочи, извержения половых продуктов при икрометании и т. д.
§24. МЕХАНИЗМ МЫШЕЧНОГО СОКРАЩЕНИЯСобытия, развивающиеся на микроскопическом (субклеточном) уровне при возбуждении мышцы, выглядят следующим образом. Потенциал действия, возникающий в результате нервной или иной стимуляции на мембране мышечного волокна, в скелетных мышцах по Т-каналам достигает мембран цитоплазматического ретикулума. Мембрана последнего высвобождает ионизированный кальций, который, в свою очередь, активизирует процесс взаимодействия актиновых и миозиновых протофибрилл.В гладких мышцах, лишенных тубулярной системы, высвобождение Са++ происходит непосредственно в самой сарколемме. А так как сарколемма напрямую контактирует с актомиозиновым комплексом гладких мышц, то активизация сокращения гладких мышц также происходит под влиянием ионизированного кальция.Интимный механизм взаимодействия актиновых и миозиновых структур до конца не расшифрован. Однако ясно, что в присутствии АТФ в поперечнополосатых мышцах при выбросе Са++ начинается скольжение тонких и толстых протофибрилл относительно друг друга (рис. 5.12). Считается, что непосредственной причиной этого движения протофибрилл является активизация актомиозиновых мостиков, которые либо изменяет угол наклона, либо подвергаются торсионному скручиванию (рис. 5.12). На поверхности воды летучая рыба достаточно продолжительно работает хвостом, развивая большую тягу, позволяющую ей преодолеть силу притяжения. Скорость полета этих мелких рыб превышает скорость движения преследователей (тунцы, меч-рыба), а пролетаемые ими расстояния достигают нескольких сотен метров.Другие виды рыб, например пальцекрыл, могут не только парить, но и выполнять сложные маневры в воздухе. Пальцекрыл поднимается к поверхности воды и скользит по ней со скоростью 18м/с. Такую высокую скорость рыба приобретает благодаря зигзагообразным движениям хвостового плавника с гипертрофированной нижней лопастью (рис. 5.15).Сильный удар хвоста поднимает рыбу в воздух на высоту 5-7 м. Пальцекрыл пролетает в воздухе до 200м, используя при этом и воздушные потоки. Рыба способна при необходимости изменить направление полета за счет движений хвостового плавника. У нее также отмечены колебательные движения грудных плавников.Скорость полета пальцекрыла сопоставима со скоростью движения современных морских судов и нередко достигает 60- 70 км/ч,Интересен механизм движений, приводящий в "боевое" положение колючки плавников таких рыб, как ерши, окуни, колюшки, сомы. Острые и крепкие колючки плавников этих рыб защищают их от хищников.В случае опасности плавники расправляются, а колючки поднимаются перпендикулярно телу. При этом несколько увеличиваются границы тела рыбы. Проглотить или схватить такую рыбу может не каждый хищник, так как надавливание на колючки не приводит к их складыванию. Следовательно, хищник при захвате неизбежно травмирует ротовую полость. Вероятнее всего, в следующий раз хищник откажется от нападения на такую рыбу.
§25. СКОРОСТЬ ДВИЖЕНИЯСкорость плавания рыбы в воде пропорциональна частоте и амплитуде колебаний тела и хвоста и укладывается в рамки определенных математических уравнений. Например, для карася, ельца и форели предельная скорость плаванияv=(1/4)[L(3f-4)],где L - длина тела; f- частота колебаний тела (хвоста).Несмотря на то что вода является более плотной средой по сравнению с воздухом, рыбам удается достичь очень высоких скоростей (км/ч):Рыба-меч 130Голубой марлин 90 Голубой тунец 80Терпуг 70Голубая акула 60Атлантический лосось 40Естественно, что для обеспечения таких скоростей природа наделила рыб рядом морфологических, гидродинамических и метаболических приспособлений.Поскольку предельные абсолютные скорости рыб зависят от их чиненных размеров, специалисты предлагают использовать для сопоставления скоростных возможностей рыб относительный показатель - коэффициент скорости:Этот коэффициент характеризует скорость рыбы, равную числу ее корпусов (длин) в секунду. Тогда все виды рыб можно классифицировать, как минимум, шестью категориями (табл. 5.4). Понятно, что рыбы с одинаковыми максимальными скоростями движения, но с различной длиной тела согласно этой классификации могут относиться к разным категориям. |
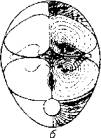 Рис. 5.8. Топография красной мускулатуры на поперечном разрезе тела тунцаФункционально красная мускулатура также выделяется. Она обеспечивает мед. ленную, но продолжительную работу. Красные мышцы меньше утомляются и быстрее восстанавливаются при интенсивных нагрузках. Именно в такого рода мышечной деятельности испытывают повышенную потребность активные пелагические рыбы - тунцы и макрели, покрывающие за сутки расстояния в сотни и тысячи километров.В качестве источника энергии красные мышцы используют жирные кислоты и кетоновые тела, что возможно лишь при наличии кислорода. В красных мышцах содержится мало гликогена и много ферментов, необходимых для аэробных реакций. Даже при предельном напряжении в красных мышцах активно протекают аэробные процессы. Поэтому так велика потребность красных мышц в кислороде и так развита в них сеть капиллярных сосудов.Мышечные волокна красной мускулатуры иннервируются медленными нервными волокнами, т. е. волокнами малого диаметра. Это частично объясняет причины большого латентного периода красных мышц. Однако на каждом мышечном волокне имеется несколько концевых пластинок, гарантирующих точность выполнения команды из центральной нервной системы.Светлые мышцы у рыб составляют основную массу активной части опорно-двигательного аппарата. Они отличаются более коротким латентным периодом (по сравнению с темными мышцами), большой скоростью сокращения и расслабления. Как для светлых, так и для красных мышц характерны тетанические сокращения, что определено природой электрического импульса, поступающего по аксону мотонейрона, - спайк с частотой 30- 50 Гц. Иннервация светлых мышц осуществляется за счет быстрых (миелинизированных) нервных волокон большого диаметра. Каждое мышечное волокно у костистых рыб имеет несколько нервно-мышечных синапсов.Кровоснабжение светлых мышц не такое интенсивное, как красных. В качестве источника энергии светлые мышцы используют глюкозу. Тот факт, что в возбужденных светлых мышцах всегда присутствует молочная кислота, свидетельствует о приоритете гликолиза в процессе мышечного сокращения.Приверженность светлых мышц к анаэробному использованию энергии глюкозы подчеркивает и высокая активность ферментов анаэробного цикла в светлых мышцах рыб. Известно, что гликолиз - неэф-фективный путь использования энергии по сравнению с окислительным фосфорилированием. В данном случае природа приносит в жертву энергетическую экономию для получения выигрыша в скорости мышечного сокращения.Клетка мышечной ткани (мышечное волокно) имеет специфическое строение. Снаружи она покрыта особой мембранной оболочкой - сарколеммой, а ее протоплазма (саркоплазма) наряду с традиционными цитозольными структурами включает в себя особые органеллы - миофибриллы,Миофибрилла является структурным элементом мышечного волокна, обеспечивающим укорочение (сокращение) (рис. 5.9).При световом микроскопировании миофибрилл видна их поперечная исчерченность из-за разницы в светопреломлении входящих в их состав белков - актина и миозина. В состав миофибрилл входят толстые и тонкие протофибриллы. Тонкие актиновые протофибриллы образуют так называемые изотропные диски (I-диски). В средней части изотропного диска расположена Z-пластина.Более толстые миозиновые протофибриллы и заходящие своими концами актиновые нити образуют анизотропные диски (А-диски). В средней части анизотропных дисков тонких нитей нет, поэтому эту зону выделяют в самостоятельную Н-зону. В области взаимного перекрытия в пределах А-диска актиновые и миозиновые нити соединены при помощи особых мостиков. Кроме того, актиновые нити закреплены на поперечных Z-пластинах (рис. 5.10).
Рис. 5.8. Топография красной мускулатуры на поперечном разрезе тела тунцаФункционально красная мускулатура также выделяется. Она обеспечивает мед. ленную, но продолжительную работу. Красные мышцы меньше утомляются и быстрее восстанавливаются при интенсивных нагрузках. Именно в такого рода мышечной деятельности испытывают повышенную потребность активные пелагические рыбы - тунцы и макрели, покрывающие за сутки расстояния в сотни и тысячи километров.В качестве источника энергии красные мышцы используют жирные кислоты и кетоновые тела, что возможно лишь при наличии кислорода. В красных мышцах содержится мало гликогена и много ферментов, необходимых для аэробных реакций. Даже при предельном напряжении в красных мышцах активно протекают аэробные процессы. Поэтому так велика потребность красных мышц в кислороде и так развита в них сеть капиллярных сосудов.Мышечные волокна красной мускулатуры иннервируются медленными нервными волокнами, т. е. волокнами малого диаметра. Это частично объясняет причины большого латентного периода красных мышц. Однако на каждом мышечном волокне имеется несколько концевых пластинок, гарантирующих точность выполнения команды из центральной нервной системы.Светлые мышцы у рыб составляют основную массу активной части опорно-двигательного аппарата. Они отличаются более коротким латентным периодом (по сравнению с темными мышцами), большой скоростью сокращения и расслабления. Как для светлых, так и для красных мышц характерны тетанические сокращения, что определено природой электрического импульса, поступающего по аксону мотонейрона, - спайк с частотой 30- 50 Гц. Иннервация светлых мышц осуществляется за счет быстрых (миелинизированных) нервных волокон большого диаметра. Каждое мышечное волокно у костистых рыб имеет несколько нервно-мышечных синапсов.Кровоснабжение светлых мышц не такое интенсивное, как красных. В качестве источника энергии светлые мышцы используют глюкозу. Тот факт, что в возбужденных светлых мышцах всегда присутствует молочная кислота, свидетельствует о приоритете гликолиза в процессе мышечного сокращения.Приверженность светлых мышц к анаэробному использованию энергии глюкозы подчеркивает и высокая активность ферментов анаэробного цикла в светлых мышцах рыб. Известно, что гликолиз - неэф-фективный путь использования энергии по сравнению с окислительным фосфорилированием. В данном случае природа приносит в жертву энергетическую экономию для получения выигрыша в скорости мышечного сокращения.Клетка мышечной ткани (мышечное волокно) имеет специфическое строение. Снаружи она покрыта особой мембранной оболочкой - сарколеммой, а ее протоплазма (саркоплазма) наряду с традиционными цитозольными структурами включает в себя особые органеллы - миофибриллы,Миофибрилла является структурным элементом мышечного волокна, обеспечивающим укорочение (сокращение) (рис. 5.9).При световом микроскопировании миофибрилл видна их поперечная исчерченность из-за разницы в светопреломлении входящих в их состав белков - актина и миозина. В состав миофибрилл входят толстые и тонкие протофибриллы. Тонкие актиновые протофибриллы образуют так называемые изотропные диски (I-диски). В средней части изотропного диска расположена Z-пластина.Более толстые миозиновые протофибриллы и заходящие своими концами актиновые нити образуют анизотропные диски (А-диски). В средней части анизотропных дисков тонких нитей нет, поэтому эту зону выделяют в самостоятельную Н-зону. В области взаимного перекрытия в пределах А-диска актиновые и миозиновые нити соединены при помощи особых мостиков. Кроме того, актиновые нити закреплены на поперечных Z-пластинах (рис. 5.10).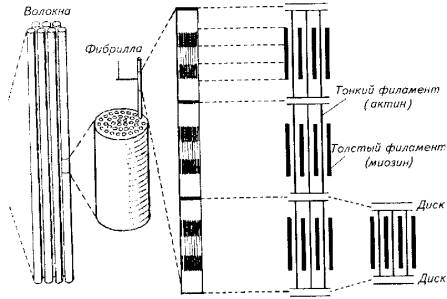 Рис. 5.9. Микроструктура скелетной мышцыИсследования позволили выявить в составе тонких актиновых нитей и элементы другого белка - миозина (в виде тропомиозина).Участок миофибриллы между двумя Z-пластинами называют саркомером.Количество и соотношение толстых и тонких протофибрилл колеблется не только у разных видов рыб, но и в разных мышцах у одной особи. Быстрые мышцы имеют большее количество нитей и меньше саркоплазмы. В медленных мышцах больше саркоплазмы.Помимо нитевидных структур - миофибрилл и протофибрилл, в составе мышечного волокна скелетных мышц выделяется хорошо развитая тубулярная система - сеть каналов и трубочек. Подсчитано, что общая поверхность мембран тубулярной системы в 10 раз превышает площадь наружной мембраны мышечного волокна. Сарколемма образует разветвленные выпячивания, проникающие в область Z-пластин. Их называют Т-канальцами. К Т-канальцам прилегает другая тубулярная система, которая относится к саркоплазматическому ретикулуму. Она образует сложную ячеистую сеть вокруг саркомера (рис. 5.11).Эта мембранная сеть обладает очень высокой ферментативной активностью. Она же выступает в роли депо ионизированного кальция, без которого невозможно мышечное сокращение.В медленных мышцах объем Т-канальцев значительно меньше по сравнению с их объемом в быстрых мышцах.Потенциал действия, возникающий на сарколемме при возбуждении мышцы, передается по Т-канальцам, а затем по каналам саркоплазматического ретикулума достигает непосредственно протофибриллярного аппарата.
Рис. 5.9. Микроструктура скелетной мышцыИсследования позволили выявить в составе тонких актиновых нитей и элементы другого белка - миозина (в виде тропомиозина).Участок миофибриллы между двумя Z-пластинами называют саркомером.Количество и соотношение толстых и тонких протофибрилл колеблется не только у разных видов рыб, но и в разных мышцах у одной особи. Быстрые мышцы имеют большее количество нитей и меньше саркоплазмы. В медленных мышцах больше саркоплазмы.Помимо нитевидных структур - миофибрилл и протофибрилл, в составе мышечного волокна скелетных мышц выделяется хорошо развитая тубулярная система - сеть каналов и трубочек. Подсчитано, что общая поверхность мембран тубулярной системы в 10 раз превышает площадь наружной мембраны мышечного волокна. Сарколемма образует разветвленные выпячивания, проникающие в область Z-пластин. Их называют Т-канальцами. К Т-канальцам прилегает другая тубулярная система, которая относится к саркоплазматическому ретикулуму. Она образует сложную ячеистую сеть вокруг саркомера (рис. 5.11).Эта мембранная сеть обладает очень высокой ферментативной активностью. Она же выступает в роли депо ионизированного кальция, без которого невозможно мышечное сокращение.В медленных мышцах объем Т-канальцев значительно меньше по сравнению с их объемом в быстрых мышцах.Потенциал действия, возникающий на сарколемме при возбуждении мышцы, передается по Т-канальцам, а затем по каналам саркоплазматического ретикулума достигает непосредственно протофибриллярного аппарата. Рис. 5.10. Взаимное расположение актиновых и миозиновых протофибрилл
Рис. 5.10. Взаимное расположение актиновых и миозиновых протофибрилл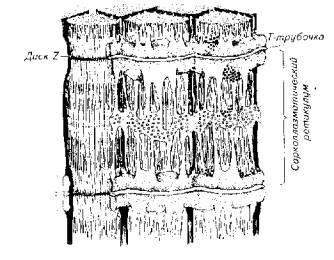 Рис. 5.11. Схема, иллюстрирующая расположение фибриллярных элементов и тубулярной сети мышечного волокнаВ целом скорость сокращения мышцы зависит от длины саркомера, количества протофибрилл и характера их расположения в пределах саркомера, количества митохондрий и энергетического материала, наличия и количества миоглобина в саркоплазме.
Рис. 5.11. Схема, иллюстрирующая расположение фибриллярных элементов и тубулярной сети мышечного волокнаВ целом скорость сокращения мышцы зависит от длины саркомера, количества протофибрилл и характера их расположения в пределах саркомера, количества митохондрий и энергетического материала, наличия и количества миоглобина в саркоплазме.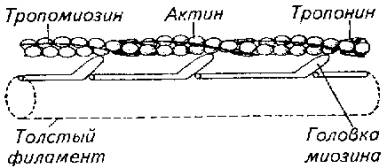 Рис. 5.12. Модель взаимодействия актиновой и миозиновой нитейУстановлена АТФ-азная зависимость конформационных изменений актомиозиновых мостиков. В отсутствие АТФ эти изменения невозможны, т. е. процесс скольжения протофибрилл не происходит.Следует подчеркнуть исключительно важную роль ионов кальция в этом процессе, который не только влияет на конформационные изменения актомиозина, но и является активатором АТФ-азы. Более того, процесс расслабления мышцы зависит от Са++. Актиновые и миозиновые нити не возвращаются в исходное положение до тех пор, пока не произойдет обратное поглощение ионов кальция мембраной цитоплазматического ретикулума или мембраной сарколеммы (в гладких мышцах), причем это обратное движение кальция зависит от активности АТФ-азы соответствующего мембранного комплекса. Поэтому не только сокращение, но и расслабление мышцы протекает с затратами энергии.Эксперименты показали, что АТФ-азную активность проявляет сам белок миозин. Он же при наличии АТФ способен соединяться с актином. Причем АТФ-азная часть миозина активируется актином. Однако белок тропонин в составе актиновой нити блокирует эту активность. И только выброс Са++ снимает тормозящее действие тропонина. Поэтому тропонин-тропомиозиновую систему рассматривают как предохранительный механизм взаимодействия актиновых и миозиновых нитей в состоянии физиологического покоя.Механизм укорочения гладкомышечного волокна менее понятен. Распространена точка зрения, согласно которой укорочение гладкомышечного волокна есть результат конформационного изменения актомиозинового комплекса. Белок актомиозин выделен из гладких мышц многих животных, включая рыб. По своим свойствам он близок к миозину млекопитающих. Как актиновая, так и миозиновая молекула имеют фибриллярную спиралевидную часть. Активизация актомиозина приводит к изменению конформации молекулы белка с соответствующим изменением его пространственного внутрицеллюлярного положения. Это и приводит к общему укорочению мышечного волокна, так как актомиозиновые структуры морфологически связаны с сарколеммой гладкомышечного волокна.Нервная регуляция гладкомышечной активности у рыб довольно специфична и заслуживает специального обсуждения. Влияние нервной системы на гладкие мышцы может быть как возбуждающим, так и тормозящим. Характер нервной стимуляции гладких мышц существенно зависит от гормонального статуса рыб и функционального состояния самого висцерального органа (например, степени наполнения желудка, мочеточников, стадии созревания половых клеток в ястыке и их наличия в матке или яйцеводе и т. д.).Гладкая мускулатура иннервируется симпатическим и парасимпатическим отделами нервной системы. Кроме того, многие висцеральные органы (например, желудочно-кишечный тракт крупные сосуды) обладают собственной интрамуральной нервной системой в виде нервных сплетений разной степени сложности.Как правило, симпатические нервные волокна в качестве медиаторов выделяют норадреналин, а парасимпатические волокна - соответственно ацетилхолин. Однако исследования показали, что в составе парасимпатической системы есть адренергические, а в составе симпатической системы -холинергические волокна. Поэтому у рыб симпатическая и парасимпатическая нервные системы являются смешанными по функциональным признакам. У ацетилхолина имеется два типа рецепторов - мускариновые и никотиновые. Для норадреналина описано также два функциональных типа рецепторов, называемых а- и р-рецепторами.Стимуляция вагуса как представителя парасимпатической части нервной системы у рыб вызывает различные эффекты не только по отношению к различным физиологическим системам, но и в пределах одной системы. Так, у форели и других желудочных рыб вагус тормозит работу желудка. Возбуждающему эффекту желудок подвергается при стимуляции чревного нерва.Далее желудка (кишечник) влияние симпатической и парасимпатической систем изменяется на прямо противоположное. Адренергические волокна и адреналин оказывают тормозящее действие, а холинергические волокна и ацетилхолин стимулируют перистальтику кишечника. Интересно и то, что ацетилхолин возбуждает все отделы кишечника у рыб. Адреналин же возбуждает только желудок и оказывает тормозящее действие на весь кишечник.У некоторых рыб пищеварительная трубка содержит элементы поперечнополосатой мускулатуры. Например, у линя кишка покрыта своеобразным чехлом подобных мышц. Поэтому его кишечник одинаково отвечает как на электрическую, так и на гуморальную стимуляцию.Кроме плавания и висцеральных функций мышцы обеспечивают и некоторые довольно специфичные движения рыб. Так, таиландский ходячий сом в составе грудных плавников имеет шипы-ходули, которыми он пользуется для переползания по суше из одного водоема в другой. Наличие таких ходуль и приспособленность к дыханию атмосферным воздухом (имеет хорошо развитую систему наджаберных полостей) позволяют этой рыбе преодолевать в период засухи довольно большие расстояния в поискахПравда, при движении по суше (точнее сказать грязи, так как рыба для передвижения все-таки выбирает увлажненные места) сом использует не только грудные плавники-ходули, но и при вытянутом теле, извиваясь подобно змее, облегчает работу мышц плавников. Эта особенность таиландского сома позволила ему освоить большие территории на юге США, куда он случайно попал из Азии.Морской петух тригла (рис. 5.13) также ползает по дну, но при этом опирается сразу на три луча грудных плавников (отсюда и его название "тригла"). Интересно, что эти ходильные лучи у петуха обособлены от других и способны сгибаться. Кроме движения по дну видоизмененные лучи грудных плавников обеспечивают этой рыбе тактильную рецепцию.В других случаях рыбы используют грудные плавники для планирования в воздухе. Так, летучие рыбы при испуге за счет броскового движения туловищных мышц, мышц хвостового стебля и интенсивной работы нижней лопастью хвостового плавника выскакивают из воды и пролетают в воздухе расстояния, по-зволяющие им избавиться от преследователей (рис. 5.14).
Рис. 5.12. Модель взаимодействия актиновой и миозиновой нитейУстановлена АТФ-азная зависимость конформационных изменений актомиозиновых мостиков. В отсутствие АТФ эти изменения невозможны, т. е. процесс скольжения протофибрилл не происходит.Следует подчеркнуть исключительно важную роль ионов кальция в этом процессе, который не только влияет на конформационные изменения актомиозина, но и является активатором АТФ-азы. Более того, процесс расслабления мышцы зависит от Са++. Актиновые и миозиновые нити не возвращаются в исходное положение до тех пор, пока не произойдет обратное поглощение ионов кальция мембраной цитоплазматического ретикулума или мембраной сарколеммы (в гладких мышцах), причем это обратное движение кальция зависит от активности АТФ-азы соответствующего мембранного комплекса. Поэтому не только сокращение, но и расслабление мышцы протекает с затратами энергии.Эксперименты показали, что АТФ-азную активность проявляет сам белок миозин. Он же при наличии АТФ способен соединяться с актином. Причем АТФ-азная часть миозина активируется актином. Однако белок тропонин в составе актиновой нити блокирует эту активность. И только выброс Са++ снимает тормозящее действие тропонина. Поэтому тропонин-тропомиозиновую систему рассматривают как предохранительный механизм взаимодействия актиновых и миозиновых нитей в состоянии физиологического покоя.Механизм укорочения гладкомышечного волокна менее понятен. Распространена точка зрения, согласно которой укорочение гладкомышечного волокна есть результат конформационного изменения актомиозинового комплекса. Белок актомиозин выделен из гладких мышц многих животных, включая рыб. По своим свойствам он близок к миозину млекопитающих. Как актиновая, так и миозиновая молекула имеют фибриллярную спиралевидную часть. Активизация актомиозина приводит к изменению конформации молекулы белка с соответствующим изменением его пространственного внутрицеллюлярного положения. Это и приводит к общему укорочению мышечного волокна, так как актомиозиновые структуры морфологически связаны с сарколеммой гладкомышечного волокна.Нервная регуляция гладкомышечной активности у рыб довольно специфична и заслуживает специального обсуждения. Влияние нервной системы на гладкие мышцы может быть как возбуждающим, так и тормозящим. Характер нервной стимуляции гладких мышц существенно зависит от гормонального статуса рыб и функционального состояния самого висцерального органа (например, степени наполнения желудка, мочеточников, стадии созревания половых клеток в ястыке и их наличия в матке или яйцеводе и т. д.).Гладкая мускулатура иннервируется симпатическим и парасимпатическим отделами нервной системы. Кроме того, многие висцеральные органы (например, желудочно-кишечный тракт крупные сосуды) обладают собственной интрамуральной нервной системой в виде нервных сплетений разной степени сложности.Как правило, симпатические нервные волокна в качестве медиаторов выделяют норадреналин, а парасимпатические волокна - соответственно ацетилхолин. Однако исследования показали, что в составе парасимпатической системы есть адренергические, а в составе симпатической системы -холинергические волокна. Поэтому у рыб симпатическая и парасимпатическая нервные системы являются смешанными по функциональным признакам. У ацетилхолина имеется два типа рецепторов - мускариновые и никотиновые. Для норадреналина описано также два функциональных типа рецепторов, называемых а- и р-рецепторами.Стимуляция вагуса как представителя парасимпатической части нервной системы у рыб вызывает различные эффекты не только по отношению к различным физиологическим системам, но и в пределах одной системы. Так, у форели и других желудочных рыб вагус тормозит работу желудка. Возбуждающему эффекту желудок подвергается при стимуляции чревного нерва.Далее желудка (кишечник) влияние симпатической и парасимпатической систем изменяется на прямо противоположное. Адренергические волокна и адреналин оказывают тормозящее действие, а холинергические волокна и ацетилхолин стимулируют перистальтику кишечника. Интересно и то, что ацетилхолин возбуждает все отделы кишечника у рыб. Адреналин же возбуждает только желудок и оказывает тормозящее действие на весь кишечник.У некоторых рыб пищеварительная трубка содержит элементы поперечнополосатой мускулатуры. Например, у линя кишка покрыта своеобразным чехлом подобных мышц. Поэтому его кишечник одинаково отвечает как на электрическую, так и на гуморальную стимуляцию.Кроме плавания и висцеральных функций мышцы обеспечивают и некоторые довольно специфичные движения рыб. Так, таиландский ходячий сом в составе грудных плавников имеет шипы-ходули, которыми он пользуется для переползания по суше из одного водоема в другой. Наличие таких ходуль и приспособленность к дыханию атмосферным воздухом (имеет хорошо развитую систему наджаберных полостей) позволяют этой рыбе преодолевать в период засухи довольно большие расстояния в поискахПравда, при движении по суше (точнее сказать грязи, так как рыба для передвижения все-таки выбирает увлажненные места) сом использует не только грудные плавники-ходули, но и при вытянутом теле, извиваясь подобно змее, облегчает работу мышц плавников. Эта особенность таиландского сома позволила ему освоить большие территории на юге США, куда он случайно попал из Азии.Морской петух тригла (рис. 5.13) также ползает по дну, но при этом опирается сразу на три луча грудных плавников (отсюда и его название "тригла"). Интересно, что эти ходильные лучи у петуха обособлены от других и способны сгибаться. Кроме движения по дну видоизмененные лучи грудных плавников обеспечивают этой рыбе тактильную рецепцию.В других случаях рыбы используют грудные плавники для планирования в воздухе. Так, летучие рыбы при испуге за счет броскового движения туловищных мышц, мышц хвостового стебля и интенсивной работы нижней лопастью хвостового плавника выскакивают из воды и пролетают в воздухе расстояния, по-зволяющие им избавиться от преследователей (рис. 5.14).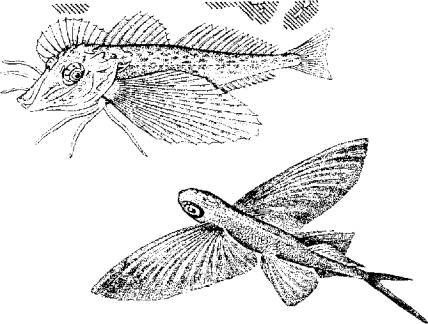 Рис. 5.13. Морской петух использует лучи грудных плавников для "хождения" по дну. Рис. 5.14. Полет пальцекрыла
Рис. 5.13. Морской петух использует лучи грудных плавников для "хождения" по дну. Рис. 5.14. Полет пальцекрыла Рис. 5.15. След на поверхности воды, оставляемый летучей рыбой при взлетеКроме того, расправленные плавники и торчащие колючки ложно увеличивают размеры рыбы, что также повышает ее шансы на выживание.Важно отметить, что колюшкам и сомам не приходится прибегать к мышечной силе, так как колючки в "боевом" положенийудерживаются не мышцами, а при помощи особого механизма. (Рис. 5.16.)B механизме фиксации колючки заложен принцип замка, который используется и другими животными (коленный сустав лошади, суставы нижних конечностей вороны и др.). Чем сильнее хищник давит на колючку, тем прочнее колючка фиксируется в замке, т е. прочность замка возрастает за счет использования энергии хищника, а не потенциальной жертвы.Опорой для колючки и ее замка служит особое образование из нескольких костей и мышц, формирующих шарнир (см. рис.5.16,///)Шарнир запирается при помощи V-образной косточки, которая имеет прочные связки с колючкой и скелетом. При возбуждении рыбы мышцы 5 и 6 сокращаются, колючка поднимается, а кость 3 блокирует шарнир. В таком положении давление на колючку делает фиксацию шарнира только жестче.Для опускания колючки необходимо, чтобы расслабились мышцы 5, 6 и 7, но сократилась мышца 8. На удержание колючки в поднятом состоянии расходуется минимум мышечных усилий, а для приведения в горизонтальное положение требуется сокращение всего одной мышцы 8.Рыбы-брызгуны используют мышечный аппарат глотки, рта и жабр для выплевывания струи воды (рис. 5.17).
Рис. 5.15. След на поверхности воды, оставляемый летучей рыбой при взлетеКроме того, расправленные плавники и торчащие колючки ложно увеличивают размеры рыбы, что также повышает ее шансы на выживание.Важно отметить, что колюшкам и сомам не приходится прибегать к мышечной силе, так как колючки в "боевом" положенийудерживаются не мышцами, а при помощи особого механизма. (Рис. 5.16.)B механизме фиксации колючки заложен принцип замка, который используется и другими животными (коленный сустав лошади, суставы нижних конечностей вороны и др.). Чем сильнее хищник давит на колючку, тем прочнее колючка фиксируется в замке, т е. прочность замка возрастает за счет использования энергии хищника, а не потенциальной жертвы.Опорой для колючки и ее замка служит особое образование из нескольких костей и мышц, формирующих шарнир (см. рис.5.16,///)Шарнир запирается при помощи V-образной косточки, которая имеет прочные связки с колючкой и скелетом. При возбуждении рыбы мышцы 5 и 6 сокращаются, колючка поднимается, а кость 3 блокирует шарнир. В таком положении давление на колючку делает фиксацию шарнира только жестче.Для опускания колючки необходимо, чтобы расслабились мышцы 5, 6 и 7, но сократилась мышца 8. На удержание колючки в поднятом состоянии расходуется минимум мышечных усилий, а для приведения в горизонтальное положение требуется сокращение всего одной мышцы 8.Рыбы-брызгуны используют мышечный аппарат глотки, рта и жабр для выплевывания струи воды (рис. 5.17).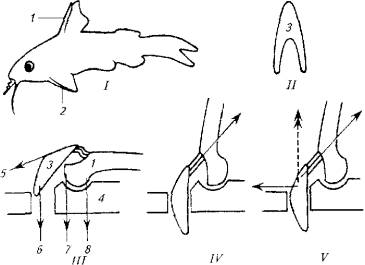 Рис. 5.16. Схема фиксации плавниковых колючек:I- общий вия рыбы; III- IV- разные положения сочленения костей; 1 - колючка спинного плавника; 2- колючка грудного плавника; 3- опорный скелет; 4, II-V-образная косточка; 5-8- места прикрепления мышц
Рис. 5.16. Схема фиксации плавниковых колючек:I- общий вия рыбы; III- IV- разные положения сочленения костей; 1 - колючка спинного плавника; 2- колючка грудного плавника; 3- опорный скелет; 4, II-V-образная косточка; 5-8- места прикрепления мышц Рис. 5.17. Рыба-брызгун: охота на насекомых при помощи струи водыРыба-брызгун достигла большого совершенства в прицельной стрельбе по насекомым, При длине собственного тела не более 20 см рыба выбрасывает мощную струю воды, которая сбивает насекомых на расстоянии до 1,5 м от поверхности воды. Впечатляет и высокая результативность такой охоты. Промахи у этой рыбы - крайне редкое явление.Все это разнообразие движений, наблюдаемое в классе рыб, обеспечивает сложная система опорно-двигательного аппарата и гладкомышечных образований, объединенных в единое целое при помощи нервной системы. Движения рыбы, в какой бы форме они ни проявлялись, осуществляют оперативное реагирование организма на изменения среды обитания с целью избежания опасности или адаптации к новым условиям.
Рис. 5.17. Рыба-брызгун: охота на насекомых при помощи струи водыРыба-брызгун достигла большого совершенства в прицельной стрельбе по насекомым, При длине собственного тела не более 20 см рыба выбрасывает мощную струю воды, которая сбивает насекомых на расстоянии до 1,5 м от поверхности воды. Впечатляет и высокая результативность такой охоты. Промахи у этой рыбы - крайне редкое явление.Все это разнообразие движений, наблюдаемое в классе рыб, обеспечивает сложная система опорно-двигательного аппарата и гладкомышечных образований, объединенных в единое целое при помощи нервной системы. Движения рыбы, в какой бы форме они ни проявлялись, осуществляют оперативное реагирование организма на изменения среды обитания с целью избежания опасности или адаптации к новым условиям.studopedya.ru
БОДИБИЛДИНГ | Вы - скульптор своего тела!
Типы волокон
Наши мышцы состоят из мышечных волокон двух видов: красных и белых. Вы знаете, что помимо расщепления креатинфосфата, мышцы используют для восстановления энергии гликолиз и окисление. Так вот, в зависимости от того, какой тип мышечных волокон работает при выполнении нагрузки, будет зависеть и способ восстановления энергии АТФ.
 Красные
волокна – это медленные волокна небольшого диаметра, которые
используют для восстановления энергии окисления. В отличие от белых волокон,
красные волокна имеют большее количество митохондрий, в которых происходит
процесс окисления для получения энергии. Красные волокна окружены мощной сетью
капилляров, необходимых для доставки большого количества кислорода с кровью.
Они не приспособлены к тяжелым и высокоинтенсивным нагрузкам, т.к. они не
способны быстро расщеплять АТФ, и получать необходимое количество энергии для
высокоинтенсивных нагрузок. Малая скорость расщепления АТФ связана в первую
очередь с низкой активностью фермента АТФазы красных волокон, с помощью,
которой как раз и происходит расщепление АТФ. То есть, к примеру, для подъема
тяжелого веса или быстрого бега красные волокна не подойдут, т.к. оба этих вида
нагрузок требуют достаточно быстрого получения и расхода энергии. Именно
поэтому красные волокна используются при незначительных нагрузках, таких как
легкий бег, ходьба или работа с легкими весами в умеренном темпе – аэробика.
Красные
волокна – это медленные волокна небольшого диаметра, которые
используют для восстановления энергии окисления. В отличие от белых волокон,
красные волокна имеют большее количество митохондрий, в которых происходит
процесс окисления для получения энергии. Красные волокна окружены мощной сетью
капилляров, необходимых для доставки большого количества кислорода с кровью.
Они не приспособлены к тяжелым и высокоинтенсивным нагрузкам, т.к. они не
способны быстро расщеплять АТФ, и получать необходимое количество энергии для
высокоинтенсивных нагрузок. Малая скорость расщепления АТФ связана в первую
очередь с низкой активностью фермента АТФазы красных волокон, с помощью,
которой как раз и происходит расщепление АТФ. То есть, к примеру, для подъема
тяжелого веса или быстрого бега красные волокна не подойдут, т.к. оба этих вида
нагрузок требуют достаточно быстрого получения и расхода энергии. Именно
поэтому красные волокна используются при незначительных нагрузках, таких как
легкий бег, ходьба или работа с легкими весами в умеренном темпе – аэробика.
Как Вы теперь знаете, что при занятиях аэробикой, кислород требуется для процессов окисления, т.е. для получения энергии.
Аэробика - это упражнения с интенсивностью 15-20% от максимальной. Работа свыше 25-30% от максимальной нагрузки, уже практически перекрывает кровообращение в нашем организме, что затрудняет доставку кислорода к мышцам и соответственно затрудняет процесс окисления, который дает энергию. Как только Вы превышаете предел аэробики, Вы практически перекрываете доставку кислорода и в работу включаются более мощные волокна – белые.
 Белые
волокна – это быстрые волокна, которые использует гликолиз, для
получения энергии. Обладающие высокой активностью фермента АТФазы, белые
волокна способны быстро расщеплять АТФ, и получать необходимую, для интенсивной
работы энергию. Но т.к. данные волокна обладают высокой скоростью расхода
энергии, они требуют и высокой скорости
восстановления молекул АТФ, которую может обеспечить только гликолиз,
т.к. в отличие от окисления он протекает непосредственно в саркоплазме мышечных
волокон, и не требует доставки кислорода митохондриям, и доставки энергии от
них уже к миофибриллам. За высокую скорость получения энергии белые волокна
платят быстрой утомляемостью, т.к. гликолиз ведет к образованию молочной
кислоты, накопление которой вызывает повышенный уровень кислотности среды мышц,
который вызывает усталость мышцы, что в конечном итоге останавливает ее работу.
В красных волокнах образование молочной кислоты не происходит, и это главная
причина их способности долго поддерживать умеренное напряжение Ваших мышц.
Белые
волокна – это быстрые волокна, которые использует гликолиз, для
получения энергии. Обладающие высокой активностью фермента АТФазы, белые
волокна способны быстро расщеплять АТФ, и получать необходимую, для интенсивной
работы энергию. Но т.к. данные волокна обладают высокой скоростью расхода
энергии, они требуют и высокой скорости
восстановления молекул АТФ, которую может обеспечить только гликолиз,
т.к. в отличие от окисления он протекает непосредственно в саркоплазме мышечных
волокон, и не требует доставки кислорода митохондриям, и доставки энергии от
них уже к миофибриллам. За высокую скорость получения энергии белые волокна
платят быстрой утомляемостью, т.к. гликолиз ведет к образованию молочной
кислоты, накопление которой вызывает повышенный уровень кислотности среды мышц,
который вызывает усталость мышцы, что в конечном итоге останавливает ее работу.
В красных волокнах образование молочной кислоты не происходит, и это главная
причина их способности долго поддерживать умеренное напряжение Ваших мышц.
Максимальную силу и максимальную скорость, которую Вы способны развивать, возможны благодаря именно белым волокнам. Работа от 25-30% и выше означает, что в ваших мышцах работают именно белые волокна.
В отличие от красных, белые волокна имеют больший диаметр, в них гораздо больше количество миофибрилл и гликогена, но меньше количество митохондрий. Большое количество гликогена в белых волокнах нужно, прежде всего, потому, что при окислении 1 молекула глюкозы заряжает 38 молекул АТФ, а при гликолизе 1 молекула глюкозы заряжает 2 молекулы АТФ. В белых волокнах находится и креатинфосфат, необходимый на начальном этапе высокоинтенсивных работ, что также вносит свой вклад в их объемы.
Существует и третий тип волокон, так называемые - Быстрые окислительно-гликолитические мышечные волокна, обладающие мощной анаэробной системой энергопродукции, приспособлены также и к выполнению достаточно интенсивной аэробной работы. С функциональной точки зрения эти волокна рассматриваются, как промежуточные между медленными и быстрыми гликолитическими. Когда совершается нагрузка при выполнении упражнений 25-40% от максимума, в работу включаются именно этот тип волокон, которые могут получать энергию, как при наличие кислорода, так и при его отсутствии (переход от работы красных волокон к белым). Когда нагрузка достигает приблизительно 40% организм уже полностью переключается на белые быстрые волокна.
В среднем человек имеет 40% красных - медленных волокон и 60% белых - быстрых волокон. Каждый человек индивидуален, поэтому в Вашем организме могут преобладать, как красные волокна, так и белые. И если Вы хотите иметь максимально развитые мышцы Вы должны уделять внимание обоим типам волокон.
Переходите к следующей главе - Управление мышечными волокнами.
bodybuild.3dn.ru